Organell
| Übergeordnet |
| Zelle |
| Gene Ontology |
|---|
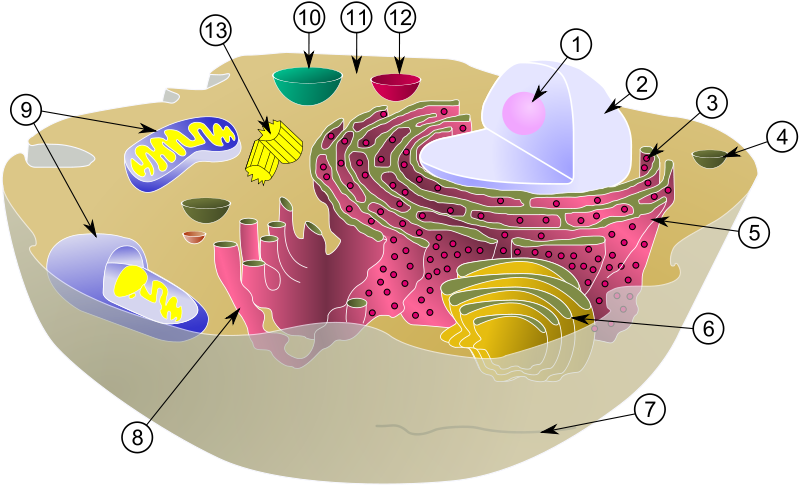
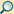
| Membranumschlossene Organellen | Strukturen ohne Membran, die je nach Definition zu den Organellen gezählt werden könnten |
|---|---|
| Zellkern (2) | Nukleolus (1) |
| Vesikel (4) | Ribosomen (3) |
| Raues Endoplasmatisches Retikulum (rER, 5) | Mikrotubuli (7) |
| Golgi-Apparat (6) | Zentriolen (13) |
| Glattes Endoplasmatisches Retikulum (sER, 8) | |
| Mitochondrien (9) | |
| Lysosom (10) | |
| Peroxisomen (12) |
Ein Organell (Diminutiv zu Organ, also „Orgänchen“) ist ein strukturell abgrenzbarer Bereich einer Zelle mit einer besonderen Funktion. Die Definition ist uneinheitlich: Manche Autoren bezeichnen nur Strukturen mit Membran als Organellen, also beispielsweise Zellkern, Mitochondrien, Plastiden (mit Chloroplasten), den Golgi-Apparat und das Endoplasmatische Retikulum. Andere fassen den Begriff weiter und schließen auch andere Strukturen ein, beispielsweise Centriolen. Bei Einzellern wird „Organell“ in diesem Sinn als Bezeichnung für komplexe Strukturen wie Geißel und Augenfleck verwendet.
Einzellige Lebewesen ohne Zellkern (Prokaryoten) haben in der Regel keine Membranen im Inneren der Zelle und demnach auch keine Organellen nach der ersten Definition. Es gibt jedoch prokaryotische Strukturen, die als Organellen im weiteren Sinn aufgefasst werden können, beispielsweise Geißeln.
Zum Auffinden weiterer zellulärer Strukturen (wie Organellen) kommen heute auch KI-gestützte Methoden zum Einsatz. Man schätzt, dass derzeit nicht einmal die Hälfte aller solcher Strukturen bekannt ist (Stand November 2021).
Begriffsgeschichte und Definitionen
Als Organ wird in der Biologie eine abgegrenzte Funktionseinheit innerhalb eines Lebewesens bezeichnet. Die Analogie zu den mikroskopischen Strukturen innerhalb einer Zelle ist für Autoren entsprechender Lehrbücher anscheinend so offensichtlich, dass sie nicht näher erläutert wird. Der erste, der für entsprechende zelluläre Strukturen eine Verkleinerungsform des Wortes ‚Organ‘ benutzte, war vermutlich der deutsche Zoologe Karl August Möbius (1884):
„Während die Fortpflanzungszellen der vielzelligen Tiere unthätig fortleben, bis sie sich loslösen, wandern und entwickeln, treten die einzelligen Tiere auch durch die an der Fortpflanzung beteiligten Leibesmasse in Verkehr mit der Außenwelt und viele bilden sich dafür auch besondere Organula.“
Organulum (Plural: Organula) ist die Verkleinerungsform zum lateinischen Organum. In einer Fußnote, die als Berichtigung in der folgenden Ausgabe der Zeitschrift erschien, erklärte Möbius:
- „Die Organe der Heteroplastiden [= Mehrzeller] bestehen aus vereinigten Zellen. Da die Organe der Monoplastiden [= Einzeller] nur verschieden ausgebildete Teile einer Zelle sind schlage ich vor, sie ‚Organula‘ zu nennen“. [die geklammerten Erklärungen kommen im Originaltext nicht vor].
Die ursprünglichste Definition des Begriffs beschränkte sich demnach ausschließlich auf Zellbestandteile von Einzellern.
Einige etwas später erschienene Arbeiten nennen Möbius namentlich als Urheber.
Es dauerte noch etliche Jahre, bis die Bezeichnung Organulum oder die neuere Entsprechung Organell sich generell durchsetzten und in einer erweiterten Bedeutung damit auch Bestandteile von Zellen mehrzelliger Organismen bezeichnet wurden. Bücher und Lehrbücher um 1900, von Valentin Häcker, Edmund Wilson und Oscar Hertwig, sprachen noch von den Organen der Zelle. Später wurden beide Bezeichnungen wohl eine Zeit lang nebeneinander verwendet: Bengt Lidforss schrieb 1915: „Eine Neubildung dieser Organe oder Organellen findet wenigstens bei höheren Pflanzen nicht statt“.
Gegen 1920 wurde der Ausdruck Organell benutzt für die Antriebsstrukturen („motor organelle complex“, Flagellen und deren Verankerung) und andere Strukturen von Einzellern. Alfred Kühn schrieb 1920 von den Centriolen als Teilungsorganellen, für welche allerdings bei den Vahlkampfien gelte, dass „die Alternative: Organell oder Produkt der Strukturbildung“ noch nicht entschieden sei – ohne aber darauf einzugehen, worin der Unterschied zwischen beiden Alternativen läge.
Max Hartmann benutzte den Ausdruck 1953 in einem Lehrbuch für extrazelluläre (Pellicula, Schalen, Zellwände) und intrazelluläre Skelette der Einzeller.
Erst später bildete sich die heute weit verbreitete Definition heraus, nach der nur von einer Membran umgebene Zellbestandteile als Organellen angesehen werden. Manchmal wird dies noch weiter eingeschränkt und nur Mitochondrien und Plastiden, die ein eigenes Genom haben, werden als Organellen bezeichnet. Aber auch die ursprünglichere Definition der subzellulären Funktionseinheiten im Allgemeinen ist weiterhin in Benutzung.
Der Ursprung der Bezeichnung Organell im deutschen Sprachraum scheint vergessen worden zu sein. Albert Frey-Wyssling schrieb 1978 vom „englischen Terminus ‚the organelle‘“, der häufig falsch mit ‚die Organelle‘ statt mit ‚das Organell‘ übersetzt würde. Frey-Wyssling schlug vor, dass sämtliche Energie verbrauchenden Strukturelemente der Zelle und nur diese als Organellen bezeichnet werden sollten, also beispielsweise auch Centrosomen, Ribosomen und Nucleoli. Diese Energie-abhängige Definition hat sich jedoch nicht durchgesetzt.
Im Gegensatz zur Bezeichnung Organell, welche sich immer auf ein einzelnes Objekt bezieht (etwa ein Mitochondrium), wird die Bezeichnung Kompartiment für die Summe aller gleichartigen zellulären Räume verwendet. Eine Zelle kann demnach viele Mitochondrien haben, aber nur ein mitochondriales Kompartiment. Auch das Cytoplasma ist ein Kompartiment, aber kein Organell.
Membranbegrenzte Organellen
Mitochondrien, der Zellkern und Plastiden (Chloroplasten und deren Verwandte) sind von einer doppelten Membran umgeben. Andere membranbegrenzte Organellen haben eine einfache Membran. Hierzu zählen die Komponenten des Endomembransystems und bei Pflanzen die Zellsaftvakuole. Daneben gibt es einige spezielle membranbegrenzte Organellen, die nur in bestimmten Zelltypen oder bestimmten eukaryotischen Artengruppen, meist Einzellern, auftreten.
Semiautonome Organellen
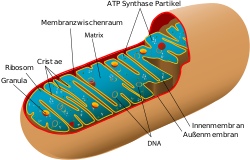
Die bei fast allen Eukaryoten vorkommenden Mitochondrien und die für Algen und höhere Pflanzen spezifischen Plastiden haben ein eigenes Genom (Mitogenom bzw. Plastom) und eine eigene Maschinerie zur Proteinbiosynthese (Mitoribosomen bzw. Plastoribosomen). Sie werden daher als ‚semiautonome Organellen‘ bezeichnet.
Nach der Endosymbiontentheorie handelt es sich bei ihnen stammesgeschichtlich gesehen um Abkömmlinge von Bakterien, die von frühen eukaryotischen Zellen aufgenommen wurden. Diese Bakterien wurden im Lauf der Evolution in die Zelle integriert. Durch die Anwesenheit der Mitochondrien-Vorläufer (von α-Proteobakterien abstammend) war es der frühen eukaryotischen, zuvor anaeroben, Zelle erstmals möglich, die sehr viel effektivere sauerstoffabhängige Energiegewinnung zu nutzen. Durch die Aufnahme von Cyanobakterien, die sich zu den Plastiden entwickelten, war die Nutzung des Sonnenlichts zur Energiegewinnung möglich: Es entstanden eukaryotische Algen und damit die Vorläufer aller Pflanzen.
Semiautonome Organellen haben eine Doppelmembran: Die äußere wird von der Wirtszelle gebildet, ist also eukaryotischen Ursprungs. Sie leitet sich ab von der bei der Aufnahme der Organell-Vorgänger abgeschnürten Plasmamembran. Die innere Membran ist prokaryotischen Ursprungs. Hierbei handelt es sich um die modifizierte Plasmamembran des Symbionten. Sie stellt eine Diffusionsbarriere für den Austausch von Molekülen und Elektronen dar.
Überflüssige Strukturen der Bakterienzellen gingen verloren, die meisten Gene wurden in den Zellkern der Wirtszelle transferiert oder gingen ebenfalls verloren. Einige Gene wurden aber auch zum Genom der Organellen zugefügt, z. B. Gene für den Austausch von Proteinen und Aminosäuren mit der Wirtszelle. Übrig blieben die heute noch vorhandenen Reste des aus einem ringförmigen DNA-Molekül bestehenden prokaryotischen Genoms und Strukturen, die für die Funktion der Organellen wichtig sind.
Semiautonome Organellen vermehren sich eigenständig durch Teilung. Bei der Teilung der Wirtszelle werden sie auf die Tochterzellen aufgeteilt.
Ebenfalls semiautonome komplexe Plastiden entstehen durch Aufnahme einer Algenzelle, deren Plastiden eine verschachtelte Struktur wie eine Matrjoschka-Puppe mit insgesamt 3 oder 4 Membranen zeigen, außen mit einer oder zwei eukaryotischen Membranen.
Andere häufige membranbegrenzte Organellen
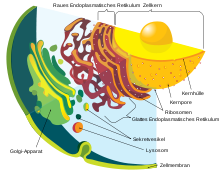
Neben den semiautonomen Organellen hat nur der Zellkern eine doppelte Membran, die Kernhülle. Die in diesem Abschnitt beschriebenen Organellen kommen in der Regel in allen Zellen eines Organismus vor. Hierzu gehören bei Pflanzen die Zellsaftvakuole und bei allen Eukaryoten verschiedene Komponenten des Endomembransystems: das Endoplasmatische Retikulum (ER), der Golgi-Apparat, Lysosomen und Peroxisomen. Eine Kurzbeschreibung dieser Organellen findet sich im Artikel Zelle an dieser Stelle. Transport-Vesikel, die für Stoffaustausch zwischen den anderen Komponenten sorgen, gehören ebenfalls zum Endomembransystem. Deren Einschluss in die Definition eines Organells ist uneinheitlich: Manchmal werden einzelne Vesikel als Organellen bezeichnet, manchmal nicht.
Tierische Zellen
| Name | Größe [μm] | Anzahl pro Zelle | % des Volumens einer Leberzelle | Funktion |
|---|---|---|---|---|
| Zellkern | 5–16 | 1 (In einem Synzytium können es mehrere sein) | 6 | Enthält die Chromosomen und damit den Hauptteil des Erbguts, Steuerzentrum der Zelle |
Endoplasmatisches Retikulum
|
? | 1 | 12 | Stoff- und Flüssigkeitstransport, Verbindungswege zwischen Zellorganellen (glatt) sowie Proteinbiosynthese (rau) |
| Golgi-Apparat | 2–3 | 1 | 3 | Bildung von Vesikeln und Lysosomen, Sekretion, Hormonbildung, |
| Mitochondrien | 0,5–1 | 1000–2000 (in einer Leberzelle) | 22 | ATP-Synthese (oxidative Phosphorylierung,, Energiegewinnung, Ort der Zellatmung, Synthese wichtiger Moleküle, Fettsäureabbau. Mitoribosomen |
| Lysosomen | 0,1–1 | 300 | 1 | Degradierung von Fremdkörpern, Autolyse nach Zelltod, intrazelluläres Recycling |
| Peroxisomen (Glyoxysomen, Microbodies) | 0,5 | 400 | 1 | Oxidierende Reaktionen (zum Beispiel zum Abbau toxischer Moleküle) |
Vesikel
|
? | 200 | 1 | Endozytose, Exozytose, Makropinozytose, intrazellulärer Transport |
Pflanzliche Zellen

In Pflanzenzellen fehlen Endosomen. Dafür haben sie Plastiden und eine Zellsaftvakuole. Eine Pflanzenzelle hat mindestens einen der Plastidtypen Chloroplast, Chromoplast und Leukoplast. Während der Differenzierung kann sich ein Plastidtyp in einen anderen umwandeln. Plastiden enthalten mit eigener DNA auch eigene Ribosomen (Plastoribosomen).
| Organell | Einzelheiten | Organellengruppe |
|---|---|---|
| Chloroplasten | Photosynthese, 2–8 μm groß | Plastiden |
| Chromoplasten | enthält Farbstoffe, zum Beispiel für Blütenfärbung, keine Photosynthese | Plastiden |
| Leukoplasten (Amyloplasten und andere) | Synthese von Monoterpenen, Aufbau und Speicherung von Stärke | Plastiden |
| Zellsaftvakuole | Speicherung von Nährstoffen, ist für den Wasserhaushalt der Zelle zuständig, Proteindegradierung u.a. Nimmt bis zu 80 % des Zellvolumens ein |
Spezielle membranbegrenzte Organellen
Zelltyp-spezifische Organellen von Mehrzellern
Die hier gelisteten Organellen kommen nur in einigen Zelltypen von bestimmten mehrzelligen Lebewesen vor, in anderen Zelltypen derselben Lebewesen aber nicht.
| Organell | Funktion | Struktur | Vorkommen |
|---|---|---|---|
| Akrosom | Hilft dem Spermium, mit dem Ei zu fusionieren | Spezielles Lysosom, von einer Membran umgeben | Spermien vieler Tiere |
| Melanosom | Farbstoffspeicher | von einer Membran umgeben | Tiere |
| Phagosom | Abbau phagozytierter Partikel | von einer Membran umgeben | Makrophagen |
Taxonspezifische Organellen
Hier sind Organellen aufgeführt, die in eukaryotischen Einzellern oder bei bestimmten mehrzelligen Arten in allen Zellen auftreten. Komplexe Plastiden wie die Apicoplasten können eigene Organellen besitzen (Nucleomorph, ein reduzierter Zellkern) und verschiedene Arten von Ribosomen. Die Plastiden der Rotalgen werden auch als Rhodoplasten, die der Glaucophyten und der photosynthetisch aktiven Arten von Paulinella (wegen ihrer Ursprünglichkeit) als Cyanellen (cyanobakterienartige Organellen) bezeichnet. Weitere komplexe Organellen sin die Augenflecken zumeist photosynthetisierender, begeißelter Protisten und vor allem die Ocelloide einzelliger Dinoflagellaten der Familie Warnowiaceae (Warnowiiden), die der Phototaxis dienen.
| Organell | Funktion | Struktur | Vorkommen | Organellgruppe |
|---|---|---|---|---|
| Ölkörper | Speicherung von Terpenen | von einer Membran umgeben | nur Lebermoose | Plastiden |
| Mitosom | Eisen-Schwefel-Cluster Assemblierung | mit Doppelmembran | einige anaerobe einzellige Eukaryoten, an Stelle von Mitochondrien. | MROs |
| Hydrogenosom | Energie- und Wasserstoffproduktion | mit Doppelmembran | einige einzellige Eukaryoten, an Stelle von Mitochondrien | MROs |
| Glycosom | Ort der Glycolyse | von einer Membran umgeben | einige Protozoen, z. B. Trypanosomen. | |
| Chromatophor | Photosynthese | von einer Membran umgeben | Paulinella chromatophora (Euglyphida) | Plastiden |
| Apicoplast | unklar, vermutlich Stoffwechselfunktionen | von vier Membranen umgeben, mit Genom | Apicomplexa, z. B. Plasmodium, Toxoplasma | Plastiden |
| MLO | unklar, vermutlich Stoffwechselfunktionen | Blastocystis | MROs | |
| Nahrungsvakuole | Aufnahme und Verdauung von Nahrung | von einer Membran umgeben | einzellige Eukaryoten | |
| Ancoracyst | Aufnahme von Nahrung | Ancoracysta | ||
| Amphoracyst (Toxicyst) | Aufnahme von Nahrung | Nebulomonas |
Erworbene temporäre Organellen
Werden semiautonome Organellen aus der Beute aufgenommen, so sind diese (per Definition) nicht auf Dauer funktionsfähig. Insbesondere sind sie in der Zelle bzw. den Zellen des Räubers nicht vermehrungsfähig und müssen daher – spätestens bei dessen Vermehrung – durch die Aufnahme (Phagocytose) weiterer Beute ersetzt werden. Handelt es sich bei den Organellen um Plastiden, spricht man von Kleptoplastidie. Bei Dinoflagellaten bleiben offenbar auch die Zellkerne der Beute eine Zeit lang funktionell (Karyokleptie). Beim Wimpertierchen Mesodinium rubrum wurde auch die Aufnahme von Beute-Mitochondrien beobachtet.
Eukaryotische Organellen ohne Membran

Eine neuere Definition setzt voraus, dass eine umgebende Membran vorhanden ist. Die ältere Bedeutung des Begriffs, die ebenfalls noch verwendet wird, kennt jedoch keine solche Voraussetzung. Nach dieser Bedeutung werden alle zellulären Strukturen, die als Organ-ähnlich angesehen werden, als Organell bezeichnet. Die Abgrenzung von derart definierten Organellen zu größeren Molekülkomplexen ist schwierig. Wenn beispielsweise Ribosomen als Organellen bezeichnet werden, warum dann nicht auch Spliceosomen oder die großen Enzymkomplexe der DNA-Replikation und Transkription? Dementsprechend ist die Zuordnung kleinerer Strukturen zu den Organellen nicht einheitlich. Bei einer Zuordnung von Ribosomen oder Nucleoli zu den Organellen würde sich ergeben, dass Organellen, und zwar Mitochondrien und Plastiden bzw. der Zellkern, selbst Organellen haben können.
Der weitere Organell-Begriff erlaubt auch den Einschluss von extrazellulären Strukturen wie der pflanzlichen Zellwände oder Schalen von Einzellern.
Eine der bekanntesten Formen der membranlosen Organellen ist das Zentrosom (auch Centrosom). Zentrosomen sind lichtmikroskopisch zu erkennen und wurden daher schon im 19. Jahrhundert entdeckt. Sie werden nicht neu gebildet, sondern entstehen durch Verdopplung und Teilung. Nach einer Zellteilung hat jede Zelle ein Zentrosom, welches sich während des Zellzyklus verdoppelt. 2006 erschien eine Arbeit, die nahelegt, dass Zentrosomen ein eigenes Genom haben. Dieses besteht nicht aus DNA, sondern aus RNA und codiert unter anderem für eine reverse Transkriptase. Sollten sich diese an der Muschel Spisula solidissima erhobenen Befunde bestätigen, müssen vielleicht auch Zentrosomen als semiautonome Organellen bezeichnet werden.
Bei der großen Vielfalt von intra- und extrazellulären Strukturen, die als Organellen gelten könnten, gibt es unter diesen Strukturen keine allgemeingültigen strukturellen oder funktionellen Gemeinsamkeiten. Die folgende, unvollständige Tabelle gibt einige Beispiele an.
| Organell | Funktion | Struktur | Vorkommen |
|---|---|---|---|
| Centrosom | Verankerung des Cytoskeletts | Zwei Centriolen und weitere Mikrotubulus-Proteine | Tiere, einige Protisten |
| Cilie | Bewegung in oder von externem Medium | Mikrotubulus-Proteine | Tiere, Protisten, einige Pflanzen |
| Myonem | Bewegung | Motorprotein-Bündel | einige Protozoen |
| Myofibrille | Muskelkontraktion | gebündelte Filamente | Tiere |
| Ribosom | Translation der mRNA in Proteine | RNA, Protein | im Cytosol aller Zellen, in Mitochondrien, Hydrogenosomen (vereinzelt), Plastiden (bis auf Ausnahmen). |
| Nucleolus | Produktion der Ribosomen | Protein, RNA, DNA | die meisten Eukaryoten |
| Vault | nicht genau bekannt | Protein, (RNA) | die meisten Eukaryoten |
| Zellwand | Stabilität | Fasern aus Zellulose oder Chitin bei Pilzen | Pflanzen, Pilze |
Prokaryotische Organellen
Nach herkömmlicher Ansicht haben Prokaryoten in der Regel in ihren Zellen keine durch Membranen abgegrenzte Bereiche und damit auch keine Organellen nach der engeren Definition. Fortschritte in der Bildgebungstechnik geben aber immer mehr Hinweise darauf, dass membranbegrenzte Organellen auch in Bakterienzellen eher die Regel und nicht die Ausnahme sind. Beispiele sind etwa Magnetosomen von magnetotaktischen Bakterien und Thylakoide der Cyanobakterien. Nach der weiter gefassten Definition des Organell-Begriffs können darüber hinaus zahlreiche weitere Strukturen so bezeichnet werden. Mesosomen, Einstülpungen der Plasmamembran von Bakterien, wurden dagegen eine Zeit lang für Organellen gehalten. Es stellte sich jedoch heraus, dass es sich um Artefakte handelte. Die folgende Tabelle listet eine Reihe dieser Organellen im engeren und in weiteren Sinn:
| Organell | Funktion | Struktur | Vorkommen |
|---|---|---|---|
| Carboxysom | Kohlenstoff-Fixierung | Schale aus Proteinen | einige Bakterien (der Cyanobakterien und Chemolithoautotrophe der Proteobakterien) |
| Metabolosom | Aldehyd-Oxidation | Schale aus Proteinen | einige Bakterien |
| Chlorosom | Photosynthese | Lichtsammelkomplex | Grüne Schwefelbakterien |
| Prokaryotische Flagelle | Bewegung | Proteinfilament | einige Prokaryoten |
| Magnetosom | Magnetische Orientierung | anorganische Kristalle, Lipidmembran | Magnetotaktische Bakterien |
| Nucleoid | DNA Aufenthaltsort, Transkription | DNA, Protein | Prokaryoten |
| Plasmid | DNA-Austausch | zirkuläre DNA | einige Bakterien |
| Ribosom | Translation der mRNA in Proteine | RNA, Protein | alle Zellen |
| Thylakoid | Photosynthese | Membran, Photosystem-Proteine und Pigmente | Cyanobakterien |
| Pepin | Kompartimentierung (DNA+Ribosomen) | Membranbläschen (Granula) | Thiomargarita magnifica |
| (Kompartiment mit Nukleoid) | Kompartimentierung (DNA+Ribosomen) | intrazytoplasmatische membranähnliche Schicht (LML) | Atribacter laminatus |
Literatur
- Karl August Möbius: Das Sterben der einzelligen und der vielzelligen Tiere. Vergleichend betrachtet. In: Biologisches Centralblatt. Band 4, Nr. 13, 14, September 1884, S. 389–392, 448 .
- Max Hartmann: Allgemeine Biologie. 4. Auflage. Gustav Fisher, Stuttgart 1953.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 26.06. 2026