Zapfen (Auge)
Zapfen oder Zapfenzelle, anatomisch Neuron coniferum (lateinisch conifer ‚Zapfen tragend‘), heißt eine Art von Fotorezeptoren in der Netzhaut des Wirbeltierauges mit kegelförmigem Fortsatz, dem Zapfen, anatomisch Conus retinae (lateinisch conus ‚Kegel‘). Zapfenzellen sind Neuronen, die als spezialisierte Sinneszellen dem photopischen Sehen bei Tageslicht dienen und für die Wahrnehmung von Farben nötig sind.
Beim Menschen werden drei Zapfentypen unterschieden, die S-, M- und L-Zapfen. Ihre Reizantwort beschreibt die spektrale Absorptionskurve, die die Grundlage der Farbmetrik bildet.
Es gibt Wirbeltiere mit einem, zwei, drei, vier oder fünf Zapfentypen.
Zapfen und Farbsehen
Als Zapfen bezeichnet man einen Typ von lichtempfindlichen Zellen in der Netzhaut des Auges. Es sind Fotorezeptoren der Wirbeltiere, die zusammen mit den Stäbchen das Sehen ermöglichen. Zapfen sind nur bei ausreichender Beleuchtungsstärke aktiv, da sie nicht sehr lichtempfindlich sind. Da bei den meisten Wirbeltieren mindestens zwei Zapfentypen unterschiedlicher spektraler Empfindlichkeit auftreten, ist ihnen in der Regel die Farbwahrnehmung möglich. Der Mensch verfügt über drei verschiedene Zapfentypen, deren Absorptionsmaxima bei etwa 455 nm, 535 nm und 563 nm liegen, diese Wellenlängen entsprechen den Farben Blauviolett, Smaragdgrün und Gelbgrün. Der mittlere Zapfentyp ist evolutiv relativ neu und verbessert die Unterscheidbarkeit von Gelb-, Rot- und Grüntönen. Die überwiegende Aktivierung bestimmter Zapfentypen ergibt entsprechende Farbtöne, ausgeglichene Reizung aller Zapfentypen ergibt dagegen die Eindrücke Grau bis Weiß. Bei übermäßiger Beleuchtungsstärke – sowohl nur in einzelnen Stellen als Glanz oder im gesamten Gesichtsfeld – sind die Zapfen überreizt (gesättigt) und es tritt Blendung ein.
Die Stäbchen sind schon bei viel geringeren Lichtintensitäten gesättigt, in Bezug auf die Helligkeit viel empfindlicher, weshalb sie im hellen Tageslicht nicht zum Seheindruck beitragen. Da beim Menschen nur ein Typ von Stäbchen vorhanden ist, kann sich durch die Stäbchen kein Farbeindruck ergeben. Bei abnehmender Lichtmenge werden die Zapfen nicht mehr ausreichend stark belichtet, dadurch scheinen die Farbtöne zu verschwinden, etwa während der Abenddämmerung. Bei der geringen Belichtungsintensität werden nur noch Stäbchen ausreichend gereizt.
Der Ausdruck „Zapfen“ wurde in historischen Kontexten anders verwendet und bezeichnete das Staphylom am Auge.
Zapfentypen
Zapfentypen des Menschen
Beim Menschen gibt es drei verschiedene Typen von Zapfen.
-
- S-Zapfen
- Short wavelength receptor oder auch K-Zapfen
(kurze Wellenlängen). Dieser
Rezeptor
deckt den blauen Bereich des sichtbaren
Farbspektrums
ab. Das Absorptionsmaximum
liegt bei einer Wellenlänge
von etwa 455 nm (blauviolett).
Ältere Quellen geben als Absorptionsmaximum 420 nm
an.
Beide Angaben liegen im Bereich blauen Lichtes, daher heißt er auch
Blaurezeptor. Der S-Zapfen des Menschen ist genetisch eng mit dem UV-Zapfen
anderer Wirbeltiere verwandt. Man spricht daher auch vom S2-Zapfen –
im Gegensatz zum S-Zapfen bei anderen Wirbeltieren.
Der Blaurezeptor ist nur selten von Farbenfehlsichtigkeit betroffen, aber wenn er nicht vorhanden ist, entsteht die Fehlsichtigkeit Tritanopie. Daher rührt die veraltete Bezeichnung T-Zapfen.
-
- M-Zapfen
- Medium wavelength receptor. Das Absorptionsmaximum des
Grünrezeptors liegt bei ungefähr 534 nm (smaragdgrün),
er deckt einen Bereich zwischen blauem und orangem Licht ab. Der menschliche
M-Zapfen ist genetisch eng mit dem L-Zapfen verwandt. Man nimmt an, dass er
sich erst vor wenigen Millionen Jahren durch eine Genduplikation aus
dem L-Zapfen entwickelt hat. L- und M-Zapfen sind auf dem X-Chromosom
nebeneinander lokalisiert. Obwohl es vier bis sechs Kopien seines Gens gibt,
ist der M-Zapfen häufig für die Farbenfehlsichtigkeit beim Menschen
verantwortlich, weil er an einer Crossing-over-Stelle
des X-Chromosoms lokalisiert ist.
Wenn keine Grünrezeptoren vorhanden sind, entsteht die Fehlsichtigkeit Deuteranopie. Daher rührt die veraltete Bezeichnung D-Zapfen.
-
- L-Zapfen
- Long wavelength receptor. Sein Absorptionsmaximum liegt
bei ungefähr 563 nm (gelbgrün).
Trotzdem wird er auch als Rotrezeptor bezeichnet, denn er übernimmt
die Hauptleistung für die Wahrnehmung von rotem
Licht. Der L-Zapfen des Menschen ist phylogenetisch
alt und entspricht dem aller anderen Wirbeltiere. Er ist auf dem X-Chromosom
kodiert.
Wenn keine Rotrezeptoren vorhanden sind, entsteht die Fehlsichtigkeit Protanopie. Daher rührt die veraltete Bezeichnung P-Zapfen.
Anzahl der Zapfentypen verschiedener Tiere
-
- 0 Zapfentypen
- Mindestens zwei nachtaktive Primatenarten besitzen keine funktionstüchtigen Zapfen; sie sehen keine Farben, sondern nur hell-dunkel.
-
- 1 Zapfentyp
- Einige Säugetiere wie Robben und die meisten Wale sind Monochromaten; sie können keine Farben unterscheiden.
-
- 2 Zapfentypen
- Die meisten Säugetiere (wie Katzen und Hunde) besitzen lediglich zwei Zapfentypen (dichromatisches Farbensehen); der M-Zapfen ist nicht vorhanden, was mit der Rot-Grün-Blindheit vergleichbar ist. Die bei tagaktiven Wirbeltieren üblichen Öltröpfchen ebenso wie die meist vorhandenen Doppelzapfen kommen noch bei Kloakentieren (Monotremen) und Beuteltieren (Marsupialiern) vor, fehlen bei Plazentatieren jedoch.
-
- 3 Zapfentypen
- Menschen, Altweltaffen und Neuweltaffen verfügen in der Regel über drei Zapfentypen; sie sehen trichromatisch.
-
- 4 Zapfentypen
- Der als ursprünglich vermutete tetrachromatische Bauplan der Wirbeltiere enthält vier verschiedene Zapfen-Typen: UV-, S-, M- und L-Zapfen, bei denen die Wellenlängen des jeweiligen Absorptionsmaximums (in der genannten Reihenfolge) um 90 bis 100 nm auseinanderliegen. Ebenso sehen viele Gliederfüßer wie Insekten und Springspinnen sowie zahlreiche Vögel tetrachromatisch. Ungefähr zwölf Prozent aller europäischen Frauen sind Tetrachromaten mit einem zusätzlichen anormalen L- oder M-Zapfen, allerdings können nur wenige dies zur genaueren Farbunterscheidung nutzen.
-
- 12 Zapfentypen
- Der Fangschreckenkrebs Neogondodactylus oerstedii besitzt acht verschiedene Rezeptortypen im sichtbaren und vier im UV-Bereich.
Spektrale Absorptionskurven
.svg.png)
Die Absorptionskurve eines Zapfentyps ist vom Bau des Opsins seines Sehpigments, dem Iodopsin, abhängig. Die photochemische Transduktion, also die Umwandlung von Lichtsignalen in neuronale Informationen, funktioniert in Stäbchen und Zapfen sehr ähnlich und in den Zapfentypen gleich. Die Reaktion eines Zapfentyps auf einen bestimmten Spektralanteil des Lichts wird daher durch seinen Opsin-Typ bestimmt.
Eine gewichtete Summe der Absorptionskurven, die die Rezeptoren bestimmen, beschreibt die Spektrale Hellempfindlichkeitskurve für das Tagsehen (die V(λ)-Kurve), ihr Maximum liegt bei 555 nm, was bei monochromatischem Licht der Farbe mittelgrün entspricht (der Wert dient auch zur Definition des Candela sowie abgeleiteter Einheiten wie Lux). Die gewichtete und für den Standardbeobachter gemittelte Summe der drei Kurven beschreibt die CIE-Tristimulus-Kurve.
Das Maximum für das Nachtsehen der Stäbchen liegt hingegen bei 498 nm (cyan).
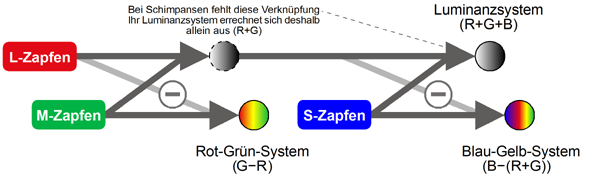
Verschaltungsmodell der Farbzellen
Die Verschaltung der Zapfen im menschlichen Auge veranschaulicht folgende Grafik:
Verteilung auf der Netzhaut
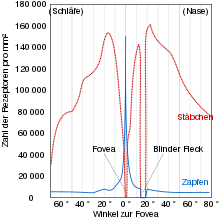
In der Fotorezeptorenschicht (Stratum neuroepitheliale) der Netzhaut des Menschen befinden sich 6 Millionen Zapfen und ungefähr 120 Millionen Stäbchen.
Der Anteil der blauempfindlichen Zapfen beträgt bei allen Menschen nahezu konstant zwölf Prozent. Das Verhältnis der Rot- und Grünzapfen auf der Retina variiert innerhalb einer Familie sehr stark. Die Dichte der Zapfen variiert zwischen den Spezies. Beim Menschen ist die Dichte der Zapfen auf der Netzhaut im Zentrum, der Fovea centralis oder „Sehgrube“, dem Bereich des schärfsten Sehens, am größten und nimmt zur Peripherie hin ab. Umgekehrt nimmt die Dichte der Stäbchen vom Zentrum zur Peripherie hin zu. Die Differenzierung in Stäbchen und Zapfen hat funktionelle Gründe: Die Zapfen funktionieren nur bei Helligkeit und Dämmerung und machen das Farbsehen möglich, während in der dunklen Dämmerung oder bei nahezu vollständiger Dunkelheit aufgrund ihrer wesentlich höheren Lichtempfindlichkeit im Wesentlichen nur noch die Stäbchen funktionieren. Die Stäbchen sind sogar in der Lage, bei absoluter Dunkelheit einzelne Photonen wahrzunehmen, wobei diese Wahrnehmung durch spontane Reaktionen auf Wärme, Augeninnendruck oder sehr starke Magnetfelder erheblich gestört werden kann.
Zellbiologischer Aufbau
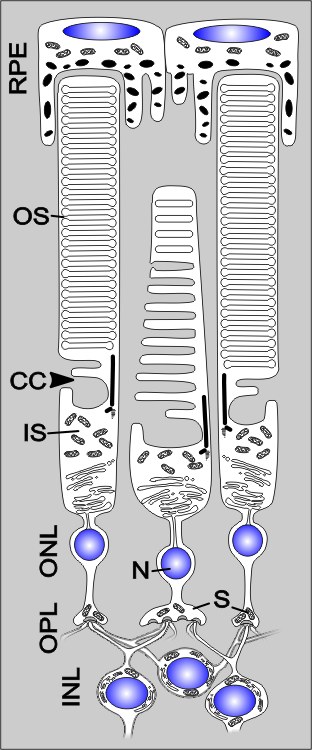
Im Aufbau sind die beiden Typen von Fotorezeptoren – Zapfen und Stäbchen – ähnlich organisiert und bestehen aus einem Zellkörper, einer Synapse sowie einer Zellspezialisierung: dem Innen- und Außensegment.
Es gibt jedoch Unterschiede:
- Die Zapfen sind wesentlich breiter als die Stäbchen.
- Bei beiden Zelltypen findet im Außensegment („Outer segment“, OS) die Phototransduktion mittels des mit Retinal gekoppelten Sieben-Transmembranproteins Opsin statt. Das Sehpigment (Opsin plus Retinal) heißt bei den Zapfen Iodopsin und ist in vielen Membraneinfaltungen eingelagert. Bei den Stäbchen befindet sich analog dazu das dem Iodopsin ähnliche Rhodopsin („Sehpurpur“) in sogenannten „Disks“.
- Die Außensegmente der Zapfen sind kürzer und müssen vom retinalen Pigmentepithel (RPE) durch verlängerte lappenartige Ausläufer kontaktiert werden, im Gegensatz zu den Stäbchen-Außensegmenten.
Ein Außensegment ist über ein modifiziertes Cilium in dezentraler Lage, das Verbindungszilium („Connecting cilium“, CC), mit dem Innensegment verbunden. Neun Mikrotubuli-Dupletts in nonagonaler Anordnung bilden die innere Struktur dieses unbeweglichen Ciliums.
An dieses schließt sich das stoffwechselaktive Innensegment („Inner segment“, IS) an. Dieses kann wiederum in das Mitochondrien-reiche Ellipsoid und in das Myoid, welches das endoplasmatische Retikulum (ER) enthält, unterteilt werden. Hier erfolgt unter anderem die Proteinbiosynthese.
Die folgende Schicht ist die äußere Körnerschicht („Outer nuclear layer“, ONL), welche den Zellkern mit dem Zellkörper beinhalten. Danach folgt die äußere plexiforme Schicht („Outer plexiform layer“, OPL) mit einer synaptischen Region. Die Synapsen am proximalen Ende der Fotorezeptoren sind teils flache, teils eingebuchtete Membranstellen. Letztere sind so genannte „Ribbon-Synapsen“, in Bezug auf eine band- oder plattenartige Struktur direkt an der aktiven Zone der Präsynapse. An die Ribbon-Struktur sind viele synaptische Vesikel gekoppelt und es können im Vergleich zu „normalen“ Synapsen eine weit höhere Anzahl von Vesikeln pro Zeiteinheit ausgeschüttet werden.
Signaltransduktion
Im Dunkeln wird von der präsynaptischen Membran des Zapfens (bzw. des Stäbchens) fortwährend der Neurotransmitter Glutamat ausgeschüttet. Trifft Licht auf den Zapfen, werden über eine Signaltransduktionskaskade Natrium-Ionenkanäle in der Zapfen-Zellmembran geschlossen. Da der Zapfen über sein Innensegment, auf Grund der dort vorherrschenden hohen Kaliumkonzentration und der dort exprimierten Kaliumionenkanäle Kalium-Ionen verliert, entwickelt er ein negatives Rezeptorpotenzial, d.h., er hyperpolarisiert und schüttet damit weniger oder kein weiteres Glutamat aus.
Der Neurotransmitter Glutamat kann erregend oder hemmend auf die nachgeschalteten Bipolarzellen wirken, denn es gibt zwei verschiedene Typen von Bipolarzellen, sogenannte ON-Bipolare und OFF-Bipolare. Verschaltet ein Zapfen auf eine ON-Bipolare, bewirkt die verminderte Ausschüttung von Glutamat bei Belichtung eine Depolarisation der ON-Bipolare. Glutamat hat eine hemmende Wirkung auf die ON-Bipolare, bei Belichtung entfällt somit die Hemmung. Dieser Effekt beruht darauf, dass in der postsynaptischen Membran der ON-Bipolare metabotrope Glutamatrezeptoren namens mGluR6 eingelagert sind. Im Dunkeln mit Glutamat besetzte mGluR6-Rezeptoren aktivieren in der ON-Bipolare eine Signalkaskade, die Kationenkanäle schließt, d.h., die Zelle wird unerregbar. Fehlt Glutamat, bleiben die mGluR6-Rezeptoren unbesetzt, die Kationenkanäle der ON-Bipolare öffnen, sie depolarisiert und leitet die Erregung weiter. Durch diesen Mechanismus wird die bei Belichtung in den Zapfen bewirkte Hyperpolarisation auf der Seite der ON-Bipolare in eine Depolarisation gewandelt, das heißt ON-Bipolare werden bei Belichtung erregt und durch Verdunkelung gehemmt.
Der zweite Typ, die OFF-Bipolaren, reagieren bei Belichtung und somit verminderter Glutamatausschüttung der Zapfen mit einer Hyperpolarisation. Sie besitzen ionotrope Glutamatrezeptoren, die unbesetzt Kationenkanäle schließen. Das heißt, OFF-Bipolare werden durch Licht gehemmt und durch Verdunkelung erregt.
Die Trennung in ON- und OFF-Bipolaren-Verschaltung bleibt in der gesamten nachfolgenden Sehbahn bis zum Gehirn erhalten. Glutamat gilt eigentlich als typischer erregender Neurotransmitter. Dieses System lässt erkennen, dass letzten Endes die postsynaptischen Glutamatrezeptoren über Erregung oder Hemmung entscheiden.
Entwicklung des Farbensehens bei Primaten
Das visuelle System der Primaten detektiert verschiedene Farben über die Aktivierung (bzw. Hyperpolarisation) der Zapfen in der Retina. Die Duplexretina des Menschens enthält drei Typen von Zapfen, deren unterschiedliche (aber überlappende) spektrale Empfindlichkeit durch drei verschiedene Opsintypen (Iodopsin) erzeugt wird. Zusammen mit einem entsprechend aufgebauten Nervensystem ermöglichen die Fotopigmente ein trichromatisches (d.h. auf der Kombination von drei Primärfarben basierendes) Farbensehen. Das Ergebnis ist die erstaunliche Fähigkeit des Menschen, rund zwei Millionen Farbnuancen unterscheiden zu können.
Die Farbwahrnehmung der Säugetiere und damit auch des Menschen, der Menschenaffen und der Primaten unterscheidet sich teils deutlich hinsichtlich der Anzahl der Sehpigmente und damit der wahrnehmbaren Farbnuancen.
Lediglich die Primaten Asiens und Afrikas (Altweltaffen), zu denen die Menschen zählen, sowie teilweise die südamerikanischen Neuweltaffen haben im Laufe ihrer Evolution die Fähigkeit des verbesserten Farbensehens erlangt. Die Fähigkeit vieler Primaten Rot von Grün zu unterscheiden, bringt mitunter lebenswichtige Vorteile mit sich. So können beispielsweise farbige reife Früchte schneller zwischen dem grünen Laub erkannt oder die jungen leichter verdaulichen Blätter von älteren schwerer verdaulichen unterschieden werden. Die nährstoffreichen Blätter weisen oftmals eine leichte Rotfärbung auf ebenso wie viele reife Früchte, die im noch grünen Stadium sogar Giftstoffe enthalten können. Deshalb setzte sich diese Fähigkeit in der Evolution durch. Bei den Altweltaffen ist eine Duplizierung eines Gens nachgewiesen worden, wodurch sich das langwelligere Sehpigment ein wenig veränderte. Dadurch enthielt das Erbgut neben der Information für das Blau-Pigment zusätzlich einen rot- und einen grün-sensitiven Sehfarbstoff. Es entstanden daraus bei vielen Primaten die drei Zapfentypen mit ihren unterschiedlichen Absorptionsmaxima.
Forschungen haben gezeigt, dass die Entwicklung des Farbensehens eng mit einer Verringerung des Geruchssinnes zusammenhängt, so ging ein Teil dieser Fähigkeit zugunsten des farbigen Sehens bei diesen Primaten verloren. Primaten mit einer besseren Geruchswahrnehmung weisen hingegen eine weniger gut ausgeprägte Fähigkeit der Farbunterscheidung auf.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 22.05. 2026