Synapse
Synapse (von griech. σύν syn ‚zusammen‘; ἅπτειν haptein ‚greifen, fassen, tasten‘) bezeichnet die Stelle einer neuronalen Verknüpfung, über die eine Nervenzelle in Kontakt zu einer anderen Zelle steht – einer Sinneszelle, Muskelzelle, Drüsenzelle oder einer anderen Nervenzelle. Synapsen dienen der Übertragung von Erregung, erlauben aber auch die Modulation der Signalübertragung, und sie vermögen darüber hinaus durch anpassende Veränderungen Information zu speichern. Die Anzahl der Synapsen beträgt im Gehirn eines Erwachsenen etwa 100 Billionen (1014) – bezogen auf ein einzelnes Neuron schwankt sie zwischen 1 und 200.000.
Der Ausdruck Synapse wurde 1897 von Charles S. Sherrington geprägt für die Verknüpfung zwischen Neuronen, beispielsweise zwischen dem aufgezweigten Ende des Axons einer Nervenzelle und dem verästeltenDendriten einer anderen Nervenzelle.
In den meisten Fällen sind es chemische Synapsen. Bei ihnen wird das Signal, das als elektrisches Aktionspotential ankommt, in ein chemisches Signal umgewandelt, in dieser Form über den zwischen den Zellen bestehenden synaptischen Spalt getragen, und dann wieder in ein elektrisches Signal umgebildet. Dabei schüttet die sendende Zelle (präsynaptisch) Botenstoffe aus, Neurotransmitter, die sich auf der anderen Seite des Spaltes (postsynaptisch) an Membranrezeptoren der empfangenden Zelle binden. Hierdurch ist die Richtung der Signalübertragung (nur vorwärts) anatomisch festgelegt, was für die Verarbeitung von Information in neuronalen Netzen grundlegend ist. Der erregungsübertragende Transmitter wird entweder in der Endigung des Axons des sendenden Neurons gebildet oder in dessen Zellkörper synthetisiert und axonal zu den präsynaptischen Membranregionen transportiert.
Dagegen sind elektrische Synapsen als gap junctions Kontaktstellen, bei denen Ionenkanäle zweier Zellen unmittelbar aneinander koppeln und so einen Übergang von Ionen und kleinen Molekülen von einer Zelle zur anderen erlauben. Zuerst wurden solche Synapsen zwischen Neuronen entdeckt, doch kommen ähnliche Kontaktstellen noch in anderen Geweben vor, auch in Pflanzen.
In übertragenem Sinn werden als immunologische Synapsen die Stellen vorübergehender zellulärer Kontakte von Zellen des Immunsystems bezeichnet, sowohl untereinander als auch mit Zellen des umgebenden Gewebes. Dabei binden Moleküle auf der Oberfläche der einen Zelle an Rezeptormoleküle und Adhäsionsmoleküle in der Zellmembran der anderen und tauschen darüber Informationen aus.
Chemische Synapsen
| Aufbau einer chemischen Synapse |
|---|
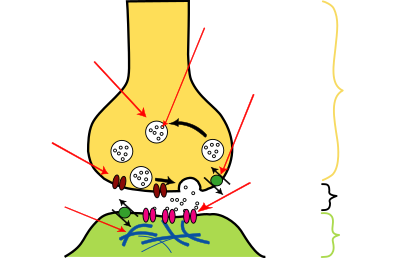
synaptisches
Bläschen Spalt
|
In einem Synapsenendknöpfchen führt das eintreffende Aktionspotential schon während der Depolarisationsphase – neben der kurzzeitigen Öffnung von Natrium- und etwas verzögert auch von Kalium-Ionenkanälen – zur vorübergehenden Öffnung spannungsaktivierter Calcium-Ionenkanäle und damit zu einem kurzdauernden Calciumioneneinstrom. Das intrazellulär erhöhte Calcium bewirkt innerhalb weniger Millisekunden die Ausschüttung eines Botenstoffs in den synaptischen Spalt. Im Endknöpfchen wird dieser Neurotransmitter in besonderen synaptischen Bläschen vorrätig gehalten und nahe der Zellmembran in synaptischen Vesikeln bereitgestellt, die unter Einwirkung von Calcium mit der präsynaptischen Membran verschmelzen können und sich dann nach außen hin entleerend so die Transmittermoleküle freisetzen.
Dieser Vorgang, der auch Exozytose genannt wird, wird erst durch die Konformationsänderung von Calcium-bindenden Proteinen möglich, insbesondere von Synaptotagminen. Sie stoßen die Bildung eines Proteinkomplexes aus SNARE-Proteinen an – aus einem Synaptobrevin in der Vesikelmembran einerseits sowie andererseits in der Zellmembran einem Syntaxin und zwei SNAP-Proteinen – der die Fusion beider Membranen erlaubt. Weitere Proteine sind dann daran beteiligt, die Öffnung des fusionierten Vesikels nach extrazellulär zu veranlassen und, wie beispielsweise Complexin I und II, die Ausschüttung der Neurotransmitter zu beschleunigen. Anschließend wird über Synapsin erneut eine bestimmte Anzahl synaptischer Vesikel am Axolemm bereitgestellt.
Auf der anderen Seite des synaptischen Spalts finden sich in der postsynaptischen, subsynaptischen, Membran der Zielzelle spezifische Rezeptormoleküle für den Neurotransmitter. Diese Rezeptoren sind zumeist mit ligandengesteuerten Ionenkanälen assoziiert (ionotrop), so dass sich unmittelbar ein Ionenkanal öffnen kann, wenn das Transmittermolekül an den passenden Rezeptor bindet. Je nach der Ionensorte, für welche dieser Kanal durchlässig ist, wird das Membranpotential in der postsynaptischen Region durch den Ionenstrom dann entweder angehoben (EPSP) oder aber abgesenkt (IPSP). Abhängig vom Rezeptortyp kann daneben mittelbar auch eine sogenannte Second-Messenger-Kaskade ausgelöst werden (metabotrop), die ebenfalls zu einer Änderung des Membranpotentials führen kann und darüber hinaus unter Umständen noch weitere Vorgänge in der postsynaptischen Zelle veranlasst. So kann – vermittelt durch den jeweiligen intrazellulären Botenstoff – auch eine Signalverstärkung hervorgerufen werden, allerdings erst mit verzögerter Wirkung.
Die Transmittermoleküle binden nicht irreversibel, sondern lösen sich nach einer gewissen Zeit wieder von ihrem Rezeptor. Im synaptischen Spalt beziehungsweise im Extrazellularraum werden sie oft durch besondere Enzyme (wie z.B. Acetylcholinesterase) abgebaut und damit in ihrer Wirkung begrenzt. Bei einigen Transmittern erfolgt kein Abbau, sondern sie werden wieder in die präsynaptische Endigung aufgenommen (beispielsweise Serotonin) oder von Gliazellen abgeräumt.
Die über chemische Synapsen übertragenen Signale haben eine biochemisch festgelegte Wirkung. Je nach Ausstattung der postsynaptischen Membran, auf die das sendende Neuron Einfluss nimmt, wird entweder eine erregende (exzitatorische) oder aber eine hemmende (inhibitorische) Wirkung erzielt. Nicht nur einzelne Synapsen, ganze Neuronen werden daher in exzitatorische und inhibitorische unterschieden, je nachdem ob sie nur erregende oder nur hemmende Synapsen an Zielzellen ausbilden. Für eine Zielzelle innerhalb des zentralen Nervensystems ist es gewöhnlich so, dass sie von verschiedenen Neuronen Signale erhält, auch gegensätzliche, und dass sich die von ihnen ausgelösten elektrischen Spannungsänderungen addieren. Überschreitet die Summe der einlaufenden exzitatorischen und inhibitorischen (postsynaptischen) Spannungsänderungen am Axonhügel dieser Nervenzelle einen bestimmten Schwellenwert bei der Potentialänderung, so wird diese Zelle ihrerseits aktiv, bildet ein Aktionspotential und leitet es über ihr Axon weiter.
Bei einer Vielzahl von psychiatrischen und neurologischen Erkrankungen wird davon ausgegangen, dass synaptische Übertragungswege gestört sind. So gibt es Anzeichen für einen Zusammenhang zwischen verschiedenen Formen von Depression und Störungen von Signalübertragungen durch den Neurotransmitter Serotonin.
Zahlreiche Medikamente oder Giftstoffe entfalten ihre Wirkung durch eine Interaktion mit Schritten der Transmission an Synapsen (Betablocker, Nicotin, Atropin, Hyoscyamin, Parathion, Kokain und viele mehr).
Elektrische Synapsen
Die Mehrzahl der Synapsen arbeitet mit einer chemischen Informationsübertragung, doch in einigen Fällen gibt es auch eine unmittelbare elektrische Weiterleitung. In diesen elektrischen Synapsen wird das Aktionspotential direkt und ohne vermittelnde Neurotransmitter an die nachfolgende Zelle weitergegeben.
Bei vielen elektrischen Synapsen findet man Verbindungskanäle durch die Zellmembran, „gap junctions“ genannt, über welche die Intrazellulärräume unmittelbar aneinander grenzender Zellen miteinander gekoppelt sind. Diese gap junctions sind Poren in der Zellmembran, die durch bestimmte Proteine, sogenannte Connexine, gebildet werden. Sechs Connexin-Moleküle kleiden dabei die Pore einer Zelle aus, zusammen bilden sie ein Connexon. Durch den Kontakt zwischen zwei Connexonen von benachbarten Zellen entsteht dann ein Kanal, der die Membranen durchquert und beide verbindet. Die offene Verbindung erlaubt eine Diffusion selbst mittelgroßer Moleküle, z.B. sekundärer Botenstoffe, und ermöglicht über Ionenpassagen eine sehr rasche Übertragung von Änderungen des Membranpotentials bei relativ geringem elektrischen Widerstand. Solche elektrischen Synapsen kommen beispielsweise zwischen Neuronen der Retina vor; sie finden sich auch zwischen Gliazellen und insbesondere zwischen Zellen des Herzmuskels, die so elektrisch zu einer gemeinsamen Einheit gekoppelt synchronisiert agieren können, ähnlich auch bei glatter Muskulatur wie dem Uterus.
Eine weitere Form der elektrischen Erregungsübertragung ist die der kapazitiven Kopplung über einen großflächigen engen Membrankontakt, wie sie beispielsweise im menschlichen Ziliarganglion zu finden ist.
Weitere Klassifikationen von Synapsen
Synapsen können des Weiteren nach verschiedenen Gesichtspunkten unterschieden werden, beispielsweise
- nach der Lage im zentralen oder im peripheren Nervensystem
- nach Art der mit einer Nervenzelle verknüpften Zelle in
- Effektorsynapsen: wirken auf einen Effektor wie Muskelzellen oder Drüsen.
- Sensorsynapsen: nehmen Signale auf von Rezeptoren wie sensiblen Sinneszellen der Haut.
- Interneuronale Synapsen: stellen Verknüpfungen zwischen Nervenzellen her und sind am häufigsten im Gehirn.
- nach den interneuronal zwischen Neuronen jeweils verknüpfenden Zellanteilen in
- Axo-dendritische Synapsen: Axonendigungen, die mit einem Dendriten des nachgeschalteten Neurons in Kontakt sind.
- Axo-somatische Synapsen: Axone oder Kollaterale, die den Zellkörper einer nachgeschalteten Nervenzelle kontaktieren.
- Axo-axonische (auch axo-axonale) Synapsen: Axon eines Neurons in Kontakt mit dem Neuriten eines anderen Neurons.
- Dendro-dendritische Synapsen: koppeln die Dendriten der Dendritenbäume verschiedener Neuronen miteinander.
- Dendro-somatische Synapsen: verknüpfen Dendriten einer Nervenzelle mit dem Körper einer anderen; so z.B. im Riechkolben.
- Somato-somatische Synapsen: verbinden den Zellkörper eines Neurons mit dem eines unmittelbar benachbarten Neurons.
- Somato-dendritische Synapsen: zwischen einem Nervenzellkörper und den Dendriten einer anderen Nervenzelle.
- Somato-axonale Synapsen: zwischen dem Zellkörper einer und dem Axon einer anderen Nervenzelle (z.B. in vegetativen Ganglien).
- nach der Weise der Zusammensetzung der miteinander verknüpften Zellen in
- komplexe Synapsen, an der mehr als zwei Zellen beteiligt sind, seriell oder parallel geschaltet.
- synaptische Glomeruli, meist von Glia umhüllte Knäuel zahlreicher Verschaltungen, konvergent oder divergent.
- reziproke Synapsen, bei der zwei Synapsen nebeneinander liegen, eine hin und eine andere her, erregend oder hemmend.
- nach der Wirkung auf die Aktivität der Zielzelle in
- exzitatorische Synapsen: erhöhen die Wahrscheinlichkeit einer Aktion (Exzitation), stimulieren oder erregen.
- inhibitorische Synapsen: verringern die Wahrscheinlichkeit einer Aktion (Inhibition), hemmen oder verhindern.
- nach der stofflichen Substanz des Neurotransmitters in
- cholinerge Synapsen
- adrenerge Synapsen
- dopaminerge Synapsen
- serotonerge Synapsen
- glutamaterge Synapsen
- glycinerge Synapsen
- GABA-erge Synapsen
- peptiderge Synapsen
Chemische Synapsen arbeiten mit unterschiedlichen Transmittern und können durch Medikamente oder Drogen in verschiedenen Schritten der Signalübermittlung verändert werden, womit je nach Angriffsort und Vorbedingungen unterschiedliche Wirkungen zu erreichen sind. Differenziertere Funktionen des Nervensystems lassen sich damit jedoch nicht gezielt beeinflussen, da diese nicht vom Überträgerstoff, sondern vom Verknüpfungsmuster der Synapsen abhängen.
Synapsengifte
Chemische Synapsengifte stören oder unterbinden die Funktion von Synapsen. Sie können die Abgabe der Neurotransmitter in den synaptischen Spalt blockieren oder den Neurotransmittern so ähnlich sein, dass sie an deren Stelle an die Rezeptormoleküle in der postsynaptischen Membran binden und damit die Erregungsübertragung stören. Je nach Bindungsweise an den Rezeptor kann damit allein ein Platz besetzt werden oder aber darüber hinaus auch eine ähnliche Wirkung erreicht werden wie durch den eigentlichen Transmitter. Nach dem erzielten Effekt werden daher Substanzen mit ähnlicher Wirkungsaktivität als Agonisten bezeichnet und unterschieden von Antagonisten mit allein der Aktivität, Agonisten in der Wirkung zu hemmen – beispielsweise indem sie deren Platz einnehmen.
Zu den bekanntesten Substanzen mit störendem Einfluss auf die synaptische Transmission gehören zahlreiche giftige Alkaloide von Pflanzen wie Atropin, Nicotin, Mescalin, Curare oder von Pilzen, etwa die des Mutterkorns oder Muskarin. Doch auch der Trinkalkohol beeinflusst die Übertragung an Synapsen, verändert z.B. GABA-Rezeptoren und blockiert (NMDA)-Glutamat-Rezeptoren. Ein schon in sehr geringer Dosis wirksames Gift ist das von einer Bakterienart der Clostridien gebildete Botulinumtoxin (Botulin) – dessen lähmende Wirkung kosmetisch zum Faltenglätten benutzt wird – und das ihm ähnliche Tetanustoxin. Zu den von Tieren gebildeten Nervengiften gehören beispielsweise die Conotoxine maritimer Kegelschnecken und die Gifte verschiedener Spinnenarten, so die Latrotoxine der dreizehnfleckigen Schwarzen Witwe. Synthetische Synapsengifte sind die chemischen Kampfstoffe Tabun, Sarin und VX und ebenso zahlreiche Insektizide, etwa E 605 oder Neonicotinoide, sowie verschiedene Halluzinogene wie LSD und andere – und selbstverständlich Psychopharmaka.
Literatur
- Susanne tom Dieck, Eckart D. Gundelfinger: Chemische Synapsen des Zentralnervensystems. Chemie in unserer Zeit 34(3), S. 140–148 (2000), ISSN 0009-2851
- Elliot Valenstein: The War of the Soups and the Sparks: The Discovery of Neurotransmitters and the Dispute Over How Nerves Communicate. 2005, ISBN 0-231-13588-2 (Buch über die Geschichte der Synapsen-Forschung)
- Gerhard Neuweiler: Die dynamische Synapse. Naturwissenschaftliche Rundschau 59(12), S. 641–650 (2006), ISSN 0028-1050
- C.S. Sherrington: The integrative action of the nervous system. Yale University Press, New Haven 1906,
- Karl-Josef Moll, Michaela Moll: Anatomie. 18. Auflage, Urban & Fischer, 2006.
- F.-J. Kretz, K. Becke: Anästhesie und Intensivmedizin bei Kindern. 2. Auflage, Georg Thieme Verlag, 2006, ISBN 978-3-13-110232-4.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 06.04. 2026