Pilze
| Pilze | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 Grüner
Anis-Trichterling (Clitocybe odora)
Grüner
Anis-Trichterling (Clitocybe odora) | ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Fungi | ||||||||||||
| L. |


Die Pilze (Fungi) bilden das dritte große Reich eukaryotischer Lebewesen neben den Tieren (Animalia) und den Pflanzen (Plantae). Sie sind wie die Pflanzen, zu denen sie lange gerechnet wurden, sesshaft, können jedoch keine Photosynthese betreiben. Daher müssen sie sich wie Tiere durch die Aufnahme organischer Substanzen ernähren (Heterotrophie), die sie jedoch in gelöster Form aus der Umgebung aufnehmen. Nach heutiger Kenntnis sind die Pilze näher mit den Tieren als mit den Pflanzen verwandt. Zu ihnen gehören vor allem Vielzeller wie die Ständerpilze aber auch Einzeller wie die Backhefe sowie coenocytische Formen mit vielen Zellkernen, aber ohne zellige Untergliederung.
Die Wissenschaft, die sich mit Pilzen beschäftigt, nennt man Mykologie.
Etymologie
Das Wort Pilz (althochdeutsch buliz) ist aus lateinisch bōlētus entlehnt; die weitere Herkunft ist unklar. Wahrscheinlich hängt das Wort mit altgriechisch βωλίτης bolites Champignon zusammen; neugriechisch βωλίτης vo̱líti̱s bezeichnet Dickröhrlinge (Boletus), insbesondere den Steinpilz (βωλίτης ο εδώδιμος), aber auch den Satans-Röhrling (βωλίτης ο σατανάς).
Die botanische Bezeichnung Fungi (lateinisch fungus ‚Pilz‘) lässt sich auf altgriechisch σφόγγος sphóngos zurückführen; dies bezeichnete ursprünglich Schwämme. Da sich Pilze ebenso mit Wasser vollsaugen wie Schwämme, wurde der Begriff im Laufe seiner Geschichte auf Pilze übertragen.
Im deutschen Sprachraum existieren die Begriffe Pilz und Schwamm oder Schwammerl parallel. Dabei wurden die Arten mit fleischiger Konsistenz als Pilze und solche, die ein festeres holz-, leder- oder korkartiges Gewebe haben, als Schwämme aufgefasst. Gleichzeitig wurde aber erkannt, dass diese Einteilung aus wissenschaftlicher Sicht nicht sinnvoll ist: Einige sehr ähnliche Arten, die derselben Gruppe angehören, wären in die beiden Kategorien aufzuteilen gewesen; außerdem hätte ein und dieselbe Art in der Jugend zu den Pilzen und im Alter zu den Schwämmen oder umgekehrt gehört. Manchmal wurden unter Schwämme auch die essbaren und unter Pilze die ungenießbaren Arten aufgefasst. Aber auch diese Einteilung ist unhaltbar. Noch heute herrscht im Südosten des deutschen Sprachraums die Bezeichnung Schwammerl vor, während sie ansonsten durch Pilz verdrängt wurde.
Mykologie, der Wortbestandteil mycetes und ähnliche Begriffe sind abgeleitet von altgriechisch μύκης mýkēs ‚Pilz‘ (Mehrzahl μύκητες mýkētes).
Vergleich mit Pflanzen und Tieren
Nachdem die Pilze wegen ihrer sesshaften Lebensweise von der Antike bis in das späte 20. Jahrhundert dem Reich der Pflanzen zugeordnet wurden, gelten sie heute aufgrund phylogenetischer, biochemischer und anatomischer Befunde als eigenes Reich und als enger mit Tieren als mit Pflanzen verwandt. Wie die Tiere gehören sie der Verwandtschaftsgruppe (Taxon) Opisthokonta an. Die Abtrennung der Pilze von den Pflanzen wurde erstmals 1969 von Robert Whittaker vorgeschlagen.
Pilze sind wie Tiere heterotroph (speziell chemoorganotroph) und ernähren sich von organischen Nährstoffen ihrer Umgebung, die sie meist durch Abgabe von Enzymen aufschließen und dadurch löslich und für sich verfügbar machen. Eine weitere Gemeinsamkeit von Pilzen und Tieren ist, dass beide das Polysaccharid Glykogen als Speichersubstanz bilden, während Pflanzen Stärke bilden. Die Abgrenzung vom Reich der Tiere erfolgt nicht aufgrund der Unbeweglichkeit der Pilze, da auch manche Tiere, wie Schwämme oder Steinkorallen, den größten Teil ihres Lebens ortsfest verbringen. Wesentliche Unterschiede zu den Tieren bestehen in der Ultrastruktur, so im Vorhandensein von Zellwänden und Vakuolen (wie bei Pflanzen).
Von den Pflanzen unterscheiden sich die Pilze vor allem durch das Fehlen von Plastiden und damit der auf Chlorophyll basierenden Photosynthese. Außerdem enthält die Zellwand der meisten Pilze neben anderen Polysacchariden auch Chitin, das im Pflanzenreich nicht vorkommt, aber der Hauptbestandteil des Exoskeletts der Gliederfüßer ist. Weiterhin fehlt den Pilzen das für Pflanzen charakteristische Polysaccharid Cellulose.
Ein grundlegender Unterschied zu den anderen höher organisierten Lebewesen ist, dass – abgesehen von der Ausbildung komplexer Strukturen wie den Fruchtkörpern – jeder Teil des Organismus autark ist und keine Kommunikation zwischen den Teilen stattfindet. Auch ein eigener Tagesrhythmus, wie ihn Tiere und Pflanzen haben, scheint bei Pilzen allenfalls als Ausnahme vorzukommen.
Gestalt und Struktur
_mycelium_on_coffee_grounds.JPG)
Der Vegetationskörper der meisten Pilze ist ein wenig differenzierter Thallus, der aus mikroskopisch feinen (2–10 µm), fädigen Hyphen besteht. Diese bilden ein weit verzweigtes Myzel, welches sich in oder auf einem festen Substrat, beispielsweise Erdboden, Holz oder anderem lebenden oder abgestorbenem organischen Gewebe, ausbreitet. Viele Pilze bilden außerdem Fruchtkörper, die sich vom Substrat abheben (z.B. die Hüte der Ständerpilze) und ebenfalls aus Hyphen bestehen (siehe unten). Daneben gibt es auch einzellige Pilze wie die Hefen.
Hyphen können durch quer liegende Septen in Zellen untergliedert sein, wobei jede Zelle einen oder zwei Kerne enthält, oder unseptiert (coenocytisch) sein und viele Kerne enthalten, die sich mit der (auch für Pflanzen charakteristischen, aber bei Metazoa nicht vorkommenden) Plasmaströmung frei bewegen können. Auch die Septen sind aber keine vollständigen Grenzen wie die Zellwände der Pflanzen, sondern haben jeweils in ihrer Mitte eine Pore (z.B. den Doliporus bei den Ständerpilzen), die einen Übergang von Cytoplasma und Organellen, darunter teils auch Kernen, ermöglicht. Daher schreiben David H. Jennings und Gernot Lysek in ihrem Buch Fungal Biology, dass die Septen keine Querwände sind und die Hyphen der Pilze grundsätzlich nicht zellig gegliedert sind, sondern ein cytoplasmatisches Kontinuum bilden. Die Septen erhöhen die Festigkeit der Hyphen; in unseptierten Hyphen finden sich stattdessen balkenartige Strukturen.
Pflanzenparasitische Pilze bilden oft Haustorien (Saugorgane) aus. Diese stülpen sich in pflanzliche Zellen, um dort Nährstoffe aufzunehmen. Einige bodenbewohnende, carnivore (fleischfressende) Pilze sind in der Lage, mit ihren Hyphen Schlingfallen für kleine Fadenwürmer (Nematoden) auszubilden (nematophage Pilze). Beim Durchkriechen werden die Nematoden dadurch festgehalten, dass sich der Hyphendurchmesser der Schlingenhyphe schnell vergrößert und sich somit die Schlingenöffnung schnell verkleinert. Eine andere Abwandlung vegetativer Hyphen sind die Substrat- oder Lufthyphen: Mehrere Bündel von Hyphen legen sich parallel aneinander und bilden makroskopisch sichtbare Hyphenstränge (Synnemata), aus denen je nach Milieu- oder Umweltänderung entweder Überdauerungsorgane (Sklerotien, Chlamydosporen) oder ungeschlechtlich erzeugte Sporen entstehen können (Konidiosporen).
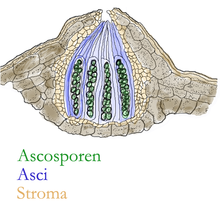
Die Fruchtkörper der Großpilze, die hut-, keulen-, knollen- oder krustenförmig sein können, bestehen aus verflochtenen Hyphen, welche ein Scheingewebe (Plektenchym) bilden. Dabei sind die Fruchtkörper nur ein kleiner Teil des gesamten Organismus und dienen der Vermehrung, Überdauerung und Ausbreitung durch Bildung von Sporen, die aus einer Meiose hervorgehen. Die Sporen werden bei vielen Pilzen in besonderen Fruchtschichten der Fruchtkörper gebildet, den Hymenien. Bei Hutpilzen befindet sich die Fruchtschicht unter dem Hut und bedeckt dort die Oberflächen der Leisten, Lamellen oder Röhren. Bei vielen Schlauchpilzen befindet sich das Hymenium knapp unter der Oberfläche des Fruchtkörpers in kleinen Kammern, den Perithezien.
Ernährung und Wachstum
Pilze ernähren sich saprotroph, indem sie gelöste Nährstoffe durch die Oberfläche ihrer Hyphen aufnehmen. Um makromolekulare, nicht lösliche Nahrungsquellen aufzuschließen, scheiden sie Enzyme aus, die den Verdauungsenzymen der Tiere (und Menschen) entsprechen.
Die Hyphen wachsen apikal (an der Spitze), ebenso wie die Wurzelhaare und Pollenschläuche der Pflanzen, aber im Unterschied zu fadenförmigen Grünalgen. Sie verzweigen sich durch seitlich aussprossende neue Spitzen, können aber auch an den Spitzen miteinander fusionieren (Anastomose) und so Netzwerke bilden.
In einem sich ausbreitenden Myzel können vier Zonen unterschieden werden:
- die Wachstumszone in der Peripherie, die aus den wachsenden Enden der Hyphen besteht,
- die sich nach innen anschließende Absorptionszone, in der Nährstoffe aus der Umgebung aufgenommen werden,
- die Speicherzone, in der ein Teil der Nährstoffe in Form von Reservestoffen abgelagert wird,
- und die zentrale Seneszenzzone, in der alte Hyphenbereiche sich dunkel verfärben und sich schließlich auflösen können.
In der Wachstumszone wandern membranumschlossene Bläschen, die als Vesikel bezeichnet werden, zur Hyphenspitze und versammeln sich dort zu dem auch lichtmikroskopisch sichtbaren Spitzenkörper. Schließlich verbinden sie sich mit der Membran an der Spitze und entlassen dabei ihren Inhalt in die sich ausdehnende Wand jenseits der Membran. Angetrieben wird das Wachstum durch die Aufnahme von Elektrolyten, insbesondere von Kalium-Ionen, und Wasser in der Absorptionszone. Dadurch wird – wie auch bei Pflanzen und Algen – der Turgor, der Druck auf die umgebende Wand, erhöht, und diese dehnt sich an der Stelle (der Hyphenspitze), wo sie dehnbar ist. Der Transport der Vesikel erfolgt entlang von Aktin-Mikrofilamenten, doch scheinen auch Mikrotubuli für die Ausrichtung der Bewegung von Bedeutung zu sein.
Neben Kalium und anderen anorganischen Elektrolyten nimmt die Hyphe im Absorptionsbereich auch lösliche Kohlenhydrate (Zucker) und Aminosäuren, die löslichen Monomere der Proteine, als Nährstoffe auf. Dies bewirkt sie durch einen Export von Protonen (H+) durch die Membran mittels eines als Protonenpumpe bezeichneten Proteins. Dadurch wird das umgebende Medium stark angesäuert, und es resultiert ein elektrochemischer Gradient. Die Enzyme für die externe „Verdauung“ (Hydrolyse) makromolekularer Nahrungsquellen werden an der Hyphenspitze ausgeschieden.
Hyphen können prinzipiell unbegrenzt weiterwachsen, so lange günstige Bedingungen vorliegen und insbesondere Nährstoffe zur Verfügung stehen. Dabei ist das Wachstum nicht chemotrop gerichtet, d.h. die Hyphen wachsen nicht in Richtung organischer Nahrungsquellen; vielmehr breitet sich das Myzel, wenn möglich, gleichmäßig in alle Richtungen aus.
Dieses trophische Wachstum endet, wenn keine Nährstoffe oder kein Sauerstoff mehr zur Verfügung steht oder wenn durch andere externe Faktoren die Fortpflanzung angeregt wird. Dies wird als Übergang von der Trophophase in die Idiophase bezeichnet. In der Idiophase werden, im Unterschied zur Trophophase, Sekundärstoffe gebildet (vgl. Sekundäre Pflanzenstoffe), die für das bloße Wachstum nicht erforderlich sind, und/oder es werden spezielle Strukturen für die Fortpflanzung ausgebildet. Das Ende der Trophophase ist für die jeweilige Hyphenspitze irreversibel.
Fortpflanzung und Vermehrung
Die meisten Pilze vermehren sich überwiegend oder ausschließlich asexuell (ungeschlechtlich). So sind bei vielen Schimmelpilzen und generell bei den Arbuskulären Mykorrhizapilzen keine sexuellen Vorgänge bekannt. Die sexuelle und überwiegend auch die asexuelle Fortpflanzung erfolgt über die Bildung und Verbreitung von Sporen. Gameten (Keimzellen) kommen nur bei den Töpfchen- oder Flagellatenpilzen vor.
Asexuelle Vermehrung
Asexuell werden die einzelligen Sporen entweder an den Enden der Hyphen abgeschnürt (Konidien), oder es werden Sporangien gebildet, in deren Innerem auf unterschiedliche Weise Sporen entstehen. Die Sporen werden dann freigesetzt, verbreiten sich und keimen schließlich zu neuen Myzelien aus. Die einzelligen Hefen vermehren sich (mit Ausnahme der Spalthefen) durch Sprossung: Nach einer Kernteilung bildet sich ein Auswuchs, in den einer der Tochterkerne einwandert und der dann abgeschnürt wird. Außerdem können die meisten Pilze sich auch durch Fragmentierung ihrer sich ausbreitenden Myzelien vermehren, weil jeder Teil des Myzels in der Lage ist, sich als eigenständiger Organismus weiterzuentwickeln.
Sexuelle Fortpflanzung
Pilze sind normalerweise haploid, haben also in ihren Zellkernen nur je einen einfachen Chromosomensatz, und durchlaufen nur bei der sexuellen Fortpflanzung eine kurze diploide Phase mit zwei Chromosomensätzen. Dazwischen liegt bei den Ständerpilzen und bei den meisten Schlauchpilzen noch eine zweikernige oder dikaryotische Phase, die bei anderen Lebewesen nicht bekannt ist. In dieser Phase enthält jede Zelle zwei haploide Kerne unterschiedlicher „elterlicher“ Herkunft. Der Ablauf der sexuellen Vorgänge unterscheidet sich bei den verschiedenen systematischen Abteilungen der Pilze sehr.
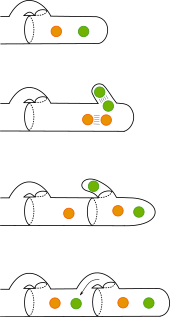
Bei den Ständerpilzen wird der Übergang von der haploiden zur diploiden Phase dadurch eingeleitet, dass zwei haploide Myzelien sich zu einem Netzwerk verbinden, indem ihre Hyphen paarweise apikal fusionieren (Anastomose). Daraus geht zunächst ein dikaryotisches Myzel hervor, in dem vor jeder Zellteilung beide Kerne sich synchron teilen und jede Tochterzelle dann zwei Kerne unterschiedlicher Herkunft erhält. Die korrekte Zuteilung der Kerne wird durch die seitliche Ausbildung einer Schnalle gewährleistet, durch welche einer der vier Kerne in eine der Tochterzellen gelangt. Das dikaryotische Myzel kann sich lange Zeit rein vegetativ ausbreiten. Die diploide Phase beginnt erst dann, wenn die beiden Kerne eines Dikaryons (einer zweikernigen Zelle) verschmelzen (Karyogamie). Das geschieht in dem als Hut aus dem Substrat herauswachsenden Fruchtkörper, wo die Enden dikaryotischer Hyphen anschwellen und sich zu den charakteristischen Basidien entwickeln, nach denen die Ständerpilze auch als Basidiomycota bezeichnet werden. In jeder Basidie entstehen durch Verschmelzung der beiden Kerne und anschließende Meiose vier haploide Zellkerne. Zugleich bildet die Basidie vier Fortsätze, in welche dann je ein Kern einwandert. Die Fortsätze werden abgeschnürt und entwickeln sich zu je einer Basidiospore, die schließlich aktiv abgeschleudert wird.
Bei den Schlauchpilzen bilden benachbarte Hyphen haploider Myzelien vielkernige sogenannte Gametangien aus, die als Ascogon und Antheridium bezeichnet werden. Das Ascogon trägt gewöhnlich eine dünne Hyphe, die Trichogyne, durch welche der Inhalt des Antheridiums in das Ascogon gelangt (Plasmogamie). Die Kerne unterschiedlicher Herkunft lagern sich eng aneinander (Kernpaarung), verschmelzen aber noch nicht miteinander. Nun wachsen aus dem Ascogon dikaryotische (oder paarkernige) Hyphen heraus. Schließlich erfolgt in der apikalen Zelle eine spezielle Zellteilung, die Hakenbildung, die der Schnallenbildung bei den Ständerpilzen ähnelt: Die Hyphenspitze krümmt sich hakenförmig zurück, die beiden Kerne teilen sich synchron, und durch Ausbildung zweier Septen resultiert eine zweikernige Tochterzelle, die jetzt an der Spitze liegt, sowie eine einkernige Stielzelle und der ebenfalls einkernige Haken. Letztere vereinigen sich dann unter Auflösung der zwischen ihnen liegenden Hyphenwände. In der jetzt apikal liegenden Zelle erfolgen nun die Karyogamie und anschließend drei Kernteilungen: eine gewöhnliche Mitose und die beiden meiotischen Teilungen (Meiose I und II). So entsteht der namengebende Schlauch oder Ascus, in dem 8 haploide Kerne in einer Reihe liegen. Anschließend werden im Ascus 8 (oder – nach weiteren Teilungen – ein Vielfaches) dickwandige Ascosporen ausgebildet und freigesetzt.
Die Jochpilze bilden keine Fruchtkörper aus, sondern existieren nur als vielkernige Myzelien. Bei ihnen senden benachbarte Hyphen als Gametangien bezeichnete Fortsätze aus, die sich zu dem namengebenden „Joch“ verbinden. Die Berührungsstelle schwillt dann an, die trennenden Zellwände lösen sich auf, und das vielkernige Verschmelzungsprodukt kapselt sich durch Trennwände von den beiden Gametangien ab. Durch paarweise Verschmelzung der Zellkerne wird die diploide Phase erreicht, und die resultierende Coenozygote (vielkernige Zygote) wird durch Ausbildung einer dicken Wand zur sogenannten Zygospore, die unter widrigen Umständen längere Zeit überdauern kann. Wenn die Zygospore unter günstigen Bedingungen auskeimt, durchlaufen die Kerne die Meiose, und es entwickelt sich wieder ein haploides vielkerniges Myzel.
Die Ausbildung von Fruchtkörpern ist mit einer erheblichen Steigerung der Stoffwechsel-Aktivität verbunden, weil in den Fruchtkörpern wesentlich mehr Proteine und Nukleinsäuren gebildet werden als im Myzel und dies einen erhöhten Energieaufwand erfordert, der sich auch in einem entsprechenden Anstieg des Sauerstoff-Verbrauchs zeigt. Deshalb können Fruchtkörper nur bei einer guten Versorgung mit Sauerstoff gebildet werden, während Myzelien rein vegetativ auch in recht sauerstoffarmen Umgebungen wie etwa in vermoderndem Holz wachsen können.
Ökologie
Pilze ernähren sich auf drei unterschiedliche Arten; als Saprophyten zersetzen sie abgestorbenes, organisches Material, oder sie ernähren sich parasitär von Lebewesen, die sie dabei beschädigen, oder sie leben als Mykorrhiza in einer wechselseitigen (mutualistischen) Symbiose im Wurzelsystem von Pflanzen oder Cyanobakterien (Flechten). Aufgrund der sehr effektiven Verbreitung ihrer Sporen sind sie praktisch überall vorhanden, wo ein geeignetes Substrat verfügbar wird, und insgesamt können sie eine sehr große Bandbreite an Nahrungsquellen nutzen.
Saprophytische Pilze

.jpg)
Die Pilze spielen eine wichtige Rolle beim Abbau organischer Materie (tote Lebewesen, Exkremente, Detritus) beteiligten Lebewesen. Die Mehrzahl der Pilze (z.B. Baumpilze) lebt saprophytisch, d.h. von organischer Substanz, die aus den Resten von toten Organismen aufgenommen werden. Gemeinsam mit Mikroorganismen und einer Reihe spezialisierter Insekten übernehmen sie die Zersetzung sämtlicher organischer Rückstände, die sie in anorganische Materie verwandeln.
Als Saprophyten leisten Pilze einen wertvollen Beitrag für den Stoffkreislauf im Boden, sowie die Anreicherung des Bodens mit Mineralstoffen, als wichtige Grundlage für Pflanzen und Kleinstlebewesen.
Zu den Stoffen, die durch diese Art von Pilzen zersetzt werden zählen Proteine, Zellulose, verschiedene Kohlenhydrate und Lignin. Komplexe Verbindungen in verholzten Zellwänden von Pflanzen enthalten Lignin und können fast ausschließlich durch saprophytische Pilze aufgespalten und verwertet werden. Auch beim Abbau von Zellulose, Hemizellulose und Keratin sind sie die wichtigsten Verwerter. Zusammen mit Bakterien und tierischen Kleinstlebewesen bilden sie aus organischem Abfall den Humus.
.jpg)
Die Bedeutung der Pilze beim Abbau des Lignins und namentlich der sehr Lignin-reichen Stämme abgestorbener Bäume ragt in mehrfacher Hinsicht heraus. Nur Pilze, und zwar speziell gewisse Ständerpilze, die als Weißfäulepilze zusammengefasst werden, sind in der Lage, größere Totholzstücke effektiv zu zersetzen. Im Unterschied zu Bakterien, von denen manche in begrenztem Maß leicht verfügbare späte Produkte des Ligninabbaus verwerten können, dringen Pilze mit ihren Hyphen aktiv in das Holz ein. Und nur darauf spezialisierte Ständerpilze verfügen über die notwendigen Enzyme für den komplizierten und energieaufwändigen Abbau des Lignins. Dieser ist unter anderem deshalb besonders schwierig, weil Lignin sehr hydrophob (wasserabstoßend) und dadurch für die gewöhnlichen hydrolytischen Abbauprozesse nicht zugänglich ist, und weil er grundsätzlich nur aerob möglich ist, also eine gute Versorgung mit Sauerstoff erfordert. Wo diese nicht gegeben ist, bleibt abgestorbenes Holz lange Zeit erhalten (etwa in Mooren) und wird schließlich über geologisch sehr lange Zeiträume in Kohle umgewandelt (Inkohlung). Von Weißfäule spricht man bei der Zersetzung von Holz durch Pilze, wenn diese vorwiegend das braune Lignin zersetzen und die farblose Zellulose übrig bleibt, während Braunfäulepilze das Lignin nur insoweit abbauen, als es für den Zugang zur Zellulose und den Hemizellulosen nötig ist.
.jpg)
Mittlerweile gibt es Hinweise, dass es auch saprophytische Pilze gibt, die sich nicht nur von toter Materie ernähren, sondern als Parasiten lebende Organismen angreifen. Ursprünglich unterschied man zwischen saprophytischen und parasitären Pilze, mittlerweile sind die Fachleute hinsichtlich der Zuordnung einiger Pilzarten unterschiedlicher Meinung. Parasiten sind auf jeden Fall auch dazu fähig, saprophytisch als Saprobionten oder Saprophyten zu leben und sich dabei auf Totholz und unbelebte Materie zu beschränken. Da die Vitalität von Bäumen in 5 Stufen bewertet wird, von 0 (gesund) bis 5 (abgestorben), sind in den Übergangsstadien bei schwer geschädigten Bäumen erhöhte Totholzanteile vorhanden. Dabei grenzen Totholzbereiche, im Baumstamm oder den Ästen an lebendes Holz, oder sind darin integriert, wobei sie noch immer einen Beitrag zus Standfestigkeit leisten. Werden solche Bereiche am lebenden Baum durch Beschädigung freigelegt, siedeln sich dort saprophytischen Holzpilzen (wie Zystidenrindenpilzen) an, die Holzfäule (z.B. Weißfäule) verursachen und die Stabilität des Baumes gefährden.
Parasitäre Pilze
Parasitär lebende Pilze sind zumeist auf bestimmte Wirtsorganismen spezialisiert, den sie angreifen und schädigen, um von seinen Nährstoffen zu profitieren, ohne dafür selbst etwas anzubieten, wie bei einer Symbiose. Um geeignete Wirte zu finden, haben diese Spezies unterschiedliche Methoden entwickelt. So produzieren Rostpilze große Mengen an Sporen und erhöhen dadurch die Chance, dass einige von ihnen auf kompatible Wirtspflanzen gelangen. Effektiver ist dagegen die Verbreitung durch Insekten, welche die Wirtspflanzen besuchen. Auf diese Weise werden etwa Hefen, die im Nektar leben, von Blüte zu Blüte transportiert. Die Sporen von Monilinia fructigena, dem Erreger der Fruchtfäule bei Obstbäumen, werden durch Wespen verbreitet, die zugleich durch Anfressen der Früchte den Zugang für den Pilz schaffen.
Brandpilze können jahrelang ohne Wirtspflanzen saprophytisch im Erdreich leben. So sind in einem von Ustilago maydis, dem Maisbeulenbrand, befallenen Acker noch bis zu 12 Jahre danach infektiöse Myzelien vorhanden, die erneut ausgesäte Maispflanzen sofort parasitieren. Auch Tiere und Menschen ziehen sich Pilzinfektionen zumeist dadurch zu, dass sie mit Sporen (Beispiel Fußpilz) oder mit anderen Wirten in Kontakt kommen.
Wenn ein Pilz eine Pflanze parasitiert, dringt er mit spezialisierten Hyphen, den Haustorien, in deren Zellen ein. Dabei durchdringen die Haustorien die Zellwand, lassen aber die Zellmembran intakt (denn andernfalls würde das Zellplasma austreten und die Wirtszelle absterben) und stülpen sie nur ein, sodass sie nun von einer Doppelmembran umgeben sind. Durch diese kann das Haustorium dann Nährstoffe aus dem Plasma der Wirtszelle entnehmen, ohne wie in toten Substraten Enzyme ausscheiden und durch fortwährendes Wachstum immer neue Nahrungsquellen erschließen zu müssen, denn die Wirtspflanze liefert die benötigten Substanzen nach, so lange die befallene Zelle am Leben bleibt.

Andererseits können Pilze die Physiologie ihrer Wirtspflanzen erheblich beeinflussen. So zeigt die Zypressen-Wolfsmilch einen stark abweichenden Habitus, wenn sie von dem Rostpilz Uromyces fabae befallen ist. Und viele Gräser sind für Weidetiere giftig, wenn sie bestimmte Pilze beherbergen. Da sie selbst dabei keine Anzeichen einer Schädigung aufweisen, kann man hier von einer mutualistischen Symbiose sprechen.
Wirtschaftlich bedeutende Pilzkrankheiten bei Pflanzen sind Maisbeulenbrand, Weizensteinbrand, Mutterkorn bei Roggen, Verticillium-Welke bei vielen Kulturpflanzen, Apfelschorf (Venturia), Birnengitterrost (Gymnosporangium sabinae), Obstbaumkrebs (Nectria galligena) und Echter Mehltau (Erysiphaceae). Daneben existieren noch circa 10.000 weitere pilzliche Pflanzenkrankheiten.
Symbiosen
Mykorrhiza

Etwa 90 Prozent aller Landpflanzen können mit bestimmten Pilzen eine Mykorrhiza bilden. Die beteiligten Pilze gehören ganz überwiegend der Klasse der Arbuskulären Mykorrhizapilze an, die mit ihren Hyphen in die Wurzelzellen eindringen (Endomykorrhiza, von gr. endo = innen) und dort durch reiche Verzweigung die namengebenden Arbuskeln (von lat. arbusculum = Bäumchen) bilden. Seltener, aber für mitteleuropäische Wälder typisch, ist die Ektomykorrhiza (von gr. ekto = außen), bei der das Pilzmyzel die Wurzeln der Bäume in Form eines Myzelmantels umschlingt und in die Rinde, nicht aber in die Zellen eindringt. Hier sind die beteiligten Pilze zumeist Ständerpilze. Wie bei jeder Symbiose profitieren beide Partner: Die Pflanze erhält über den Pilz mehr mineralische Nährstoffe, da sein feines Myzel den Boden enger durchwirkt, als ihre eigenen Saugwurzeln das könnten. Diese bessere Versorgung macht sich insbesondere in sehr nährstoffarmen Böden bemerkbar. Umgekehrt erhält der Pilz Zucker, den die Pflanze durch Photosynthese erzeugt, als Energiequelle und für die Bildung anderer organischer Substanzen. Daneben sind Mykorrhizapilze allerdings vielfach auch in der Lage, saprophytisch organische Nährstoffe aus dem Erdreich zu gewinnen.

Einen Extremfall stellen die Orchideen dar, von denen viele schon bei der Keimung ihrer Samen unter natürlichen Bedingungen obligat auf ihre pilzlichen Symbiosepartner angewiesen sind. Manche Orchideen, z.B. die Vogel-Nestwurz, enthalten kein Chlorophyll und können daher keine Photosynthese treiben, sondern beziehen alle Nährstoffe von dem Pilz, auf dem sie somit parasitieren. Die gleichen Verhältnisse finden sich auch bei manchen Heidekrautgewächsen wie dem Fichtenspargel. In beiden Fällen bilden die beteiligten Pilze zugleich eine Mykorrhiza mit Bäumen und beziehen von diesen Zucker, wovon sie einen Teil an die Nestwurz bzw. den Fichtenspargel weitergeben (Epiparasitismus). Auf diese Weise können diese Pflanzen in Form von blassen Blütenständen auch an schattigen Stellen im Wald gedeihen.
Flechten
Flechten sind Pilze, die einzellige Grünalgen oder Cyanobakterien als Symbionten beherbergen und dadurch photoautotroph, d.h. dank der Photosynthese ihrer Symbionten nicht auf externe Nahrungsquellen angewiesen sind. Sie können, ganz anders als jeder der Partner allein, extreme Lebensräume besiedeln. Dabei sind die betreffenden Pilze ohne ihre jeweiligen Symbionten kaum lebensfähig, während letztere auch isoliert gedeihen. Für sie liegt der Vorteil der Symbiose darin, dass sie ihnen ein viel breiteres Spektrum an Lebensräumen eröffnet.
Marine und xerophile Pilze
Auch in marinen Lebensräumen, also in stark salzhaltigem Milieu, sind Pilze, insbesondere Schlauchpilze, verbreitet. Dem hohen osmotischen Druck begegnen sie durch eine entsprechende Anreicherung von Polyolen (höherwertigen Alkoholen), hauptsächlich Glyzerin, aber auch Mannit und Arabit, in den Hyphen. Ähnlich verhält es sich bei xerophilen Schimmelpilzen und Hefen, die etwa auf Salzheringen oder auf Marmelade wachsen können.
Anaerobie: Leben ohne Sauerstoff
Die allermeisten Pilze benötigen Sauerstoff; sie sind obligat aerob. Manche können jedoch zeitweilig ohne Sauerstoff auskommen (fakultative Anaerobie) oder haben sogar die Fähigkeit verloren, ihn überhaupt zu nutzen (obligate Anaerobie). Letzteres trifft auf die Neocallimastigaceae zu, die im Pansen von Wiederkäuern leben und auf die Verwertung von Zellulose spezialisiert sind. Fakultative Anaerobier sind dagegen die Hefen, die unter anaeroben Bedingungen zur Gärung übergehen, mit der sie – wesentlich weniger effektiv als mit der aeroben Atmung – z.B. von Zucker leben können. Auch manche Schimmelpilze sind dazu in der Lage, z.T. gehen sie dann auch morphologisch in ein hefeartiges Stadium über.
Reaktion auf Licht
Myzelien wachsen zumeist im Dunkeln. Wenn Hyphenspitzen die dem Licht ausgesetzte Oberfläche des Substrats erreichen, regt das Licht (genauer: dessen blaue Anteile) die Bildung von Sporen an, und auch die Entwicklung der Fruchtkörper kann lichtabhängig sein. Dieser Effekt ist jedoch lokal begrenzt und wirkt sich nicht auf das übrige Myzel aus. Bei vielen Pilzen wachsen die sporenbildenden Hyphen (Konidiophoren oder Sporangiophoren) in die Richtung des einfallenden Lichtes. Bei der Gattung Pilobolus (Mucorales) wird schließlich das gesamte Sporangium, das die reifen Sporen enthält, exakt in Richtung der Lichtquelle abgeschleudert.
Verbreitung
Pilze sind sehr weit verbreitet. Die große Mehrheit ist landlebend. Im Wasser lebende, also aquatische Pilze, sind beispielsweise unter den Chytridiomycetes zu finden. Süßwasser- sind häufiger als Salzwasserpilze.
Kommunikation
Eine 2022 veröffentlichte Studie untersuchte die elektrischen Impulse der Hyphen einiger Pilzarten (des Australischen Geisterpilzes, des Samtfußrüblings, des Spaltblättlings sowie der Puppen-Kernkeule). Die Phasen der elektrischen Aktivität waren artspezifisch und dauerten von einer bis 21 Stunden an, mit einer Spannung von maximal 0,03 bis 2,1 mV. Der Autor stellt Ähnlichkeiten der Spannungsmuster mit der Struktur der menschlichen Sprache fest. Einige Pilzarten verwenden demnach bis zu 50 „Wörter“, davon 15–20 häufiger. Die komplexeste „Satzstruktur“ der untersuchten Arten verwendet der Spaltblättling. Analog zu neuronalen Netzwerken oder der Kommunikation von Bäumen mithilfe ihres Wurzelwerks könnten auch Pilze sich mit diesen elektrischen Impulsen miteinander verständigen.
Bedeutung für den Menschen
Etwa 180 Pilzarten können beim Menschen verschiedene Pilzkrankheiten hervorrufen. Weit größer ist aber der Nutzen vieler Pilze für den Menschen, etwa als Speisepilze oder bei der Herstellung von Hefeteig und alkoholischen Getränken.
Speise- und Giftpilze

Viele Pilzarten sind bekannte und beliebte Nahrungsmittel. Dazu gehören nicht kultivierbare Arten wie Steinpilz und Pfifferling, aber auch Kulturarten und -sorten von Champignon, Shiitake und Austernpilz. Beim Sammeln von Wildpilzen ist größte Sorgfalt geboten, um nicht durch versehentlich geerntete Giftpilze eine Pilzvergiftung zu riskieren. Wer Pilze für den Verzehr sammelt, muss unbedingt die Speise- und Giftpilze gründlich kennen und darf nur zweifelsfrei erkannte Speisepilze nehmen. Viele Pilzarten enthalten Hämolysine oder andere hitzelabile Gifte, die erst durch Erhitzen zerstört werden. Die meisten Speisepilze erfordern daher Erhitzen durch Kochen oder Braten vor dem Verzehr, um Verdauungsbeschwerden oder Vergiftungen zu vermeiden.
Zudem ist zu beachten, dass Pilze Schwermetalle und Radionuklide aufnehmen und anreichern. Dies kann zu gesundheitsgefährdenden Konzentrationen von Schwermetallen beziehungsweise Radionukliden im Fruchtkörper von Wildpilzen führen.
UV-exponierte Pilze können erhebliche Mengen an Vitamin D enthalten.
Die meisten Speisepilze gehören zu den Ständerpilzen. Relativ wenige Speisepilzarten, darunter die Morcheln und die Trüffeln, stammen aus der Abteilung der Schlauchpilze.
Bei manchen Pilzen unterscheidet sich der Speisewert in verschiedenen Regionen. Einige Arten wie beispielsweise der Wollige Milchling, die gemeinhin als ungenießbar gelten, werden in Osteuropa für Speisezwecke verwendet. Selbst giftige Arten wie die Frühjahrslorchel werden in Skandinavien verzehrt. Auch in derselben Region kann sich die Einstufung der Genießbarkeit innerhalb mehrerer Jahrzehnte ändern. Beispielsweise galt der heute als giftig angesehene Kahle Krempling früher als essbar.
Alkoholische Getränke, Hefeteig und Milchprodukte
Von den einzelligen Pilzen sind die Zuckerhefen der Gattung Saccharomyces, insbesondere die Backhefe (S. cerevisiae), die bekanntesten Nutzpilze. Sie erzeugen durch alkoholische Gärung aus Zucker Alkohol und Kohlendioxid und werden in der Bierbrauerei, bei der Herstellung von Wein, sonstiger alkoholischer Getränke und bestimmter (auch alkoholischer) Sauermilchprodukte sowie zum Backen verwendet. In der Regel verwendet man heute Reinzuchthefen, doch insbesondere bei der Weinherstellung werden vielfach weiterhin die natürlicherweise auf der Oberfläche der Weinbeeren lebenden Hefen verwendet. Der beim Brotbacken verwendete Sauerteig enthält neben Milchsäurebakterien auch Hefe.
Bei der Weinherstellung spielt außerdem der Myzelpilz Botrytis cinerea eine Rolle. Er erzeugt bei herbstlich kühlfeuchtem Wetter bei den Beeren eine Edelfäule, die bewirkt, dass die Beerenhaut perforiert wird. Der dadurch bedingte Wasserverlust erhöht die Zuckerkonzentration.
Viele Arten spielen auch beim Reifeprozess von Milchprodukten, insbesondere von Sauermilchprodukten und Käse, eine bedeutende Rolle.
Medizinisch bedeutende Pilze
Seit Beginn des 20. Jahrhunderts nutzt man Pilze auch für medizinische Zwecke. Medikamente wie das Antibiotikum Penicillin werden aus Pilzen gewonnen. Weitere Stoffwechselprodukte von Pilzen wirken cholesterinsenkend oder helfen gegen Malaria.
Andererseits verursachen Pilze bei Menschen Erkrankungen. Die am häufigsten betroffenen Körperstellen sind die Haut (insbesondere an Kopf, Füßen und Händen), Haare, Nägel und Schleimhäute. Die wohl bekanntesten Pilzkrankheiten des Menschen sind Haut- und Nagelpilzerkrankungen.
Auf der Haut des Menschen lebt eine Vielzahl von Bakterien und Pilzen, die ihm aber normalerweise nicht schaden. Sie siedeln in den oberen Hautschichten und ernähren sich von abgestorbenen Hautzellen und Schweiß. Faktoren wie Stress, ein geschwächtes Immunsystem, hormonale Umstellungen o.Ä. können dazu führen, dass ansonsten harmlose Pilze Krankheiten auslösen, die die Kopfhaut, die Scheide (bei einer beginnenden Schwangerschaft) oder andere innere Organe befallen. Beispiele für einen Befall innerer Organe sind Hefen wie Candida und der Schimmelpilz Aspergillus fumigatus, der vor allem nach einer Chemotherapie gelegentlich zu Erkrankungen der Lunge führt.
Fußpilze sind weit verbreitet, da sie sehr leicht übertragen werden. Einige ihrer Sporen überleben jahrelang und sind gegen normale Hygienemaßnahmen unempfindlich. Weiterhin werden sie sehr leicht von den Füßen auf andere Körperstellen wie Geschlechtsorgane, Mund und Schleimhäute übertragen. Schwimmbäder gehören zu den Hauptquellen von Fußpilzen.
Weitere Beispiele sind:
- Malassezia furfur, der Erreger von Pityriasis versicolor, einer häufigen Hautmykose
- Candida albicans, ein meist harmloser Mitbewohner, der bei Abwehrschwäche zur Erkrankung führen kann
- Aspergillus-Arten, zum Beispiel A. fumigatus als der häufigste Erreger der Aspergillose, einer Lungenerkrankung
- Cryptococcus neoformans, der Erreger der Kryptokokkose
- Rhizopus, eine Gattung der Mucorales, Erreger der Mucormykose
- Coccidioides immitis, der vor allem in den Südstaaten der USA, in Mexiko und Argentinien die Kokzidioidomykose hervorruft
- Histoplasma capsulatum, ein Endoparasit des retikuloendothelialen Gewebes und Erreger der Histoplasmose
Medikamente zur Behandlung von Pilzkrankheiten werden Antimykotika genannt. Sie werden bei lokalem Pilzbefall von Haut oder Schleimhäuten und auch bei systemischen Pilzinfektionen angewendet.
In China sind zahlreiche Großpilze seit Jahrhunderten Bestandteil der traditionellen chinesischen Medizin. Der Shiitake (Lentinula edodes) galt schon in der Ming-Dynastie (1368–1644) als Lebenselixier, das Erkältungen heilen, die Durchblutung anregen und die Ausdauer fördern sollte. Der Glänzende Lackporling (Ganoderma lucidum), bekannt als Ling-Zhi oder Reishi, soll ein besonders wirksames Tonikum sein. Der Pom-Pom-Pilz oder Igel-Stachelbart/Affenkopfpilz (Hericium erinaceus) wird bei Erkrankungen des Magens empfohlen. Der europäische Apothekerschwamm oder Lärchenbaumschwamm (Laricifomes officinalis) ist als Heilmittel hoch geschätzt. Sein wirksamer Bestandteil ist Agaricinsäure, die stark abführend wirkt und für den außerordentlich bitteren Geschmack verantwortlich ist.
Psychoaktive Pilze
Als psychoaktive Pilze oder Rauschpilze werden Pilze bezeichnet, die psychotrope Stoffe wie Psilocybin, Psilocin, Baeocystin, Muscimol oder Ergin enthalten. Am bekanntesten sind psilocybinhaltige Pilze, die oftmals als Magic Mushrooms bezeichnet werden. Ihre Wirkung wird gelegentlich mit der Rauschwirkung von LSD verglichen. Zu ihnen gehören exotische Arten wie der Kubanische (Psilocybe cubensis) oder der Mexikanische Kahlkopf (Psilocybe mexicana), aber auch einheimische Arten wie der Spitzkegelige Kahlkopf (Psilocybe semilanceata). Unerfahrene Pilzsucher riskieren mit dem Sammeln psilocybinhaltiger Pilze ihre Gesundheit aufgrund der Verwechslungsgefahr mit anderen, giftigen Pilzarten. Der Fliegenpilz enthält die giftige und selbst schon psychotrope Ibotensäure, die beim Trocknen in das wesentlich wirksamere Alkaloid Muscimol umgewandelt wird; beide Substanzen werden den Delirantia zugerechnet. Das Mutterkorn beinhaltet neben anderen (giftigen) Stoffen auch das psychoaktive Ergin. Psychoaktive Pilze hatten und haben noch heute bei verschiedenen Völkern eine spirituelle Bedeutung als entheogene Stoffe.
Zunderschwamm

Der als Baumschädling vor allem in Buchen und Birken wachsende Zunderschwamm, Fomes fomentarius, ein Weißfäulepilz, wurde früher zum Feuermachen verwendet: Das Innere der aus den Baumstämmen konsolartig herauswachsenden Fruchtkörper wurde gekocht, getrocknet, weichgeklopft, mit Salpeterlösung getränkt und erneut getrocknet. Der so erhaltene Zunder kann durch Funken entzündet werden.
Durch bloßes Kochen, Trocknen und Weichklopfen kann aus dem Fruchtkörper-Inneren auch ein dem Filz ähnliches Material gewonnen werden, das zur Herstellung verschiedener Gebrauchsgegenstände (Mützen, Taschen und dergleichen, siehe Bild) verwendet werden kann.
Pilzfarbstoffe
Die Chemie der Pilzfarbstoffe ist wegen der Vielzahl der Verbindungen sehr komplex. Einige Farbstoffe liegen in reduzierter Form als Leukoverbindung vor. So wird der Farbstoff Atromentin, ein Terphenylchinon, des Samtfußkremplings bei Verletzung des Pilzes mit Luftsauerstoff und im Pilz vorhandener Oxidasen zu einer blauen Form oxidiert. Ähnliches kann man beim Anschneiden vieler Pilze beobachten. Farbstoffe vom Pulvinsäuretyp kommen bei Dickröhrlingsverwandten, insbesondere der Gattungen Boletus und Xerocomus, vor. Der rote Farbstoff von Hexen-Röhrlingen ist Variegatorubin, der gelbe Farbstoff des Gold-Röhrlings ein Gemisch von Grevillin B und C. Grevilline sind als Farbstoffe bei den Schmierlingen von Bedeutung. Die Huthaut des Fliegenpilzes enthält zahlreiche gelbe, orange und rote Komponenten, die zur Gruppe der Betalaine gehören, sowie Muscaflavin, das auch für die orangen bzw. roten Farben von Saftlingen verantwortlich ist. Im Strubbelkopfröhrling konnte man L-Dopa nachweisen, das bei Verletzung des Fruchtkörpers unter Melaninbildung zu einer Schwarzfärbung führt. Bei bestimmten Pilzen gilt die Biosynthese von Betalaminsäure, die mit Aminosäuren Betalaine bildet, aus L-Dopa als gesichert. Darüber hinaus kommen in Pilzen häufig Carotinoide, Azulenderivate, Anthrachinone, Phenoxazine und Riboflavin vor.
Systematik
Äußere Systematik
| ||||||||||||||||||||||||||||||||||||||||||
| Äußere Systematik der Pilze |
Die nächsten Verwandten der Pilze sind neben den Holozoa (zu denen auch die vielzelligen Tiere (Metazoa) und deren Schwestergruppe, die Kragengeißeltierchen (Choanomonada) gehören) vor allem die parasitisch lebende Gattung Rozella. Ob auch die einzelligen Mikrosporidien (Microsporidia, auch Microspora genannt) zu den Pilzen zu zählen sind, ist derzeit noch unklar. Die kleinste gemeinsame Klade von Pilzen und Tieren wird als Opisthokonta bezeichnet und nach Adl et al. 2012 folgendermaßen aufgestellt:
- Opisthokonta
- Holozoa (u.a. Vielzellige Tiere und Kragengeißeltierchen)
- Nucletmycea (u.a. Pilze und Rozella)
Als gemeinsamer Vorfahr von Tieren und Pilzen kann ein geißeltragender Einzeller (Flagellat) angenommen werden, der biologisch demnach sowohl den heutigen Töpfchenpilzen als auch den Kragengeißeltierchen (Choanoflagellata) ähnelte.
Aufgrund der im Laufe der Zeit erweiterten Erkenntnisse zur Systematik werden einige Taxa, die früher als Niedere Pilze bezeichnet wurden, inzwischen nicht mehr zu den Pilzen gezählt. Dies trifft auf die Schleimpilze und andere pilzähnliche Protisten wie die Eipilze (Oomycota), die Netzschleimpilze (Labyrinthulomycetes), Hyphochytriales (einzige Ordnung der Hyphochytriomycota), die früher Plasmodiophoromycetes genannten Phytomyxea und die früher den Phycomycetes (hingegen heute den Ichthyosporea) zugerechneten Taxa Ichthyophonae (auch Amoebidiidae genannt) und Eccrinales zu.
Innere Systematik
| |||||||||||||||||||||||||||||||||||||||||||||
| Innere Systematik der Pilze |
Man kennt heute etwa 120.000 Pilzarten. Nach aktuellen Schätzungen existieren aber zwischen 2,2 und 5,1 Millionen Arten. Jedes Jahr werden etwa 1500 neue Arten beschrieben. Die früher auch Echte Pilze oder Höhere Pilze (Eumycota) genannten Lebensformen werden in die folgenden fünf Abteilungen unterteilt:
- Töpfchenpilze (Chytridiomycota) sind meist einzellige Pilze. Weil begeißelte Stadien vorhanden sind, werden die Töpfchenpilze als sehr ursprüngliche Form der Pilze angesehen.
- Jochpilze (Zygomycota) unterscheiden sich von den anderen Pilzen durch die Bildung der namensgebenden jochartigen Brücken zwischen kompatiblen Hyphen während der sexuellen Fortpflanzung. Das Myzel ist meist vielkernig (coenocytisch) ohne Septen. Die Jochpilze bilden wahrscheinlich keine natürliche Verwandtschaftsgruppe.
- Die arbuskulären Mykorrhizapilze (Glomeromycota) sind ebenfalls coenocytisch und bilden eine typische Endomykorrhiza aus, bei der bäumchenartige Membranausstülpungen, die Arbuskel, in das Innere von pflanzlichen Wurzelzellen wachsen und auf diese Weise eine symbiotische Beziehung etablieren.
- Die Zellen der Schlauchpilze (Ascomycota) sind durch Septen voneinander getrennt und enthalten meist nur einen Zellkern. Die geschlechtlichen Sporen werden in charakteristischen Schläuchen, den Asci gebildet. Es gibt eine Reihe von Arten, bei denen große Fruchtkörper auftreten und die man daher den Großpilzen zurechnet.
- Auch die Zellen der Ständerpilze (Basidiomycota) sind durch Septen voneinander getrennt, enthalten aber während einer oft ausgedehnten Phase ihrer Entwicklung (Dikaryophase) zwei verschiedene Zellkerne. Die geschlechtlichen Sporen werden an Basidien gebildet. Die meisten Großpilzarten entstammen dieser Gruppe. Das Myzel kann in Extremfällen wie beim Hallimasch mehrere tausend Jahre alt werden.
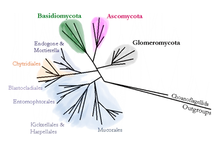
Technische Fortschritte in der molekularen Genetik und die Anwendung von computerunterstützten Analysemethoden haben detaillierte und sichere Aussagen über die systematischen Beziehungen der oben aufgeführten Pilztaxa zueinander ermöglicht. Beispielsweise wurden manche Verwandtschaften bestätigt, die vorher aufgrund morphologischer, anatomischer und physiologischer Unterschiede oder Gemeinsamkeiten nur vermutet werden konnten.
Die Töpfchenpilze haben sich demnach sehr früh von den anderen Pilzen abgespalten und viele ursprüngliche Merkmale, wie begeißelte Sporen, bewahrt. Die Jochpilze dagegen stellen sehr wahrscheinlich keine einheitliche (monophyletische) Verwandtschaftsgruppe, sondern eine polyphyletische Gruppe verschiedener Abstammungslinien dar. Die Gattung Amoebidium, die bisher zu den Jochpilzen gezählt wurde, gehört demnach nicht einmal zum Pilzreich. Die arbuskulären Mykorrhizapilze, die ursprünglich ebenfalls zu den Jochpilzen gestellt wurden, werden heute als eigenständige Verwandtschaftsgruppe angesehen, die meist in den Rang einer eigenen Abteilung erhoben wird. Sie wird dann als evolutionäre Schwestergruppe eines Taxons aus Schlauch- und Ständerpilzen angesehen, das man als Dikaryomycota bezeichnet.
Diejenigen Arten, die vorläufig nicht eindeutig einer der oben genannten Gruppen zugeordnet werden können, wurden provisorisch zu den Fungi imperfecti (Deuteromycota) gestellt; dies stellte jedoch nur ein provisorisches und künstliches Formtaxon dar.

Im Frühjahr 2007 veröffentlichten 67 Wissenschaftler aus 13 Ländern als Teil des Projekts Assembling the Fungal Tree of Life das abschließende Resultat einer konzertierten und umfassenden Forschungsanstrengung mit dem Ziel, die bislang inkonsistente und unklare Taxonomie der Pilze zu bereinigen. Dabei berücksichtigten sie nicht nur neueste molekulare und genetische Daten verschiedener Pilzspezies, sondern auch die Entwicklungsgeschichte der jeweiligen Nomenklatur. Als Ergebnis schlagen die Forscher eine neue Klassifizierung vor, in der das Reich der Pilze in 195 Taxa untergliedert wird. Auf diese Weise hoffen die Taxonomen, den bestehenden Bezeichnungswirrwarr in der wissenschaftlichen Literatur zu beenden und eine durchgehende Konsistenz der verschiedenen (Online-)Datenbanken zu erreichen.
Ein Beispiel der Veränderungen in der derzeitigen Systematik der Pilze, die sich daraus ergeben, ist die Auflösung des Phylums der Jochpilze (Zygomycota), zu denen auch bestimmte auf Früchten lebende Schimmelpilze gehören. Die betroffenen Taxa würden auf andere Gruppen aufgeteilt werden.
Evolution
Der gemeinsame Vorfahre der Pilze und der Tiere war wahrscheinlich ein begeißelter Einzeller, der vor rund einer Milliarde Jahren gelebt hat. Während die große Mehrheit der heute lebenden Pilze keine Geißeln mehr ausbildet, treten bei der Fortpflanzung der Flagellatenpilze noch begeißelte Sporen oder Gameten auf. Wahrscheinlich verließen die Pilze schon vor den Pflanzen das Wasser und besiedelten das Land. Da die frühesten bekannten Landpflanzen noch keine richtigen Wurzeln hatten, aber offenbar in Symbiose mit Arbuskulären Mykorrhizapilzen lebten, nimmt man an, dass der Landgang der Pflanzen erst durch diese Pilze ermöglicht wurde.
Fossilien

Die frühesten Fossilien mit für Pilze typischen Merkmalen stammen aus der Zeit des Paläoproterozoikums vor etwa 2,4 Milliarden Jahren. Diese mehrzelligen benthischen Organismen hatten filamentöse Strukturen, die zur Anastomose fähig waren.
Die ersten weitgehend unumstrittenen Pilzfunde stammen aus der erdgeschichtlichen Epoche des Ordoviziums und können vielleicht den Arbuskulären Mykorrhizapilzen zugeordnet werden. Der erfolgreiche Landgang der Pflanzen wäre ohne Pilzsymbiosen vermutlich nicht möglich gewesen.
Der älteste beschriebene Pilz ist Gondwanagaricites magnificus. Es handelt sich um einen fossilen Lamellenpilz aus Brasilien, der ungewöhnlich gut erhalten ist und etwa 5 cm Durchmesser hat. Der Fund stammt aus der Unterkreide und ist 115 Millionen Jahre alt.
Fossile Pilze sind ferner aus Bernsteinfunden u.a. auf karbonischer Lagerstätte in Schottland und England (sogenannter Middletonit), aus dem Karnium (Obertrias) in Deutschland und in bemerkenswerter Artenvielfalt aus kreidezeitlichem kanadischen Bernstein sowie dem mexikanischen, dominikanischen und baltischen Bernstein (alle Tertiär) bekannt. Bei einigen dieser Funde handelt es sich um Pilze, die Termiten und Nematoden befallen hatten und zusammen mit ihren Wirten vom Harz eingeschlossen wurden.
Die ältesten bekannten Fossilien fleischfressender Pilze sind etwa 100 Millionen Jahre alt (Grenze zwischen Ober- und Unterkreide). Sie wurden von Forschern der Humboldt-Universität zu Berlin um Alexander Schmidt in Bernstein aus dem Südwesten Frankreichs gefunden. Die Art lebte im küstennahen Wald und bildete wohl eine Übergangsform zwischen hefeähnlichen aquatischen Pilzen und modernen fleischfressenden Pilzen.
Kulturgeschichte

Der griechische Arzt Pedanios Dioscurides schrieb schon im ersten Jahrhundert nach Christus in seinem Lehrbuch davon, dass es zwei Arten von Schwämmen gebe: Die einen sind zum Essen bequem, die anderen aber ein tödlich Gift. Dioscurides vermutete (fälschlich), dass die Giftigkeit eines Pilzes von seinem Standort abhänge: Pilze, die neben verrostetem Eisen, faulendem Tuch, Schlangenhöhlen oder Bäumen mit giftigen Früchten wachsen, seien alle miteinander giftig. Er erkannte aber schon damals die schwere Verdaulichkeit von übermäßiger Speisepilz-Kost, die den Menschen würgen und ersticken ließen. Auch Adamus Lonicerus schrieb im 16. Jahrhundert in seinem Kräuterbuch über die Pilze, dass es die Natur aller Schwämme sei, zu bedrängen; sie seien kalter, phlegmatischer, feuchter und roher Natur.
Auch später und teilweise bis heute haben sich einige Vermutungen über Anhaltspunkte gehalten, die zur Unterscheidung essbarer und giftiger Pilze dienen sollen. Eine der bekanntesten ist der Trugschluss, dass Fruchtkörper, die von Tieren angefressen wurden, nicht giftig seien. Diese Annahme entspricht der Vorstellung, dass Pilze, die für Tiere unschädlich sind, auch für Menschen ungiftig seien. Weitere vermeintliche Indikatoren sind, dass Pilze, die bei Schlangennestern, Schimmelstellen oder giftigen Bäumen wachsen, giftig seien oder dass bei Berührung mit Gift Löffel aus Zinn oder Silber braun anlaufen, Zwiebeln sich schwarz färben, Eiweiß bleigrau oder Salz gelb. Spätestens seit Mitte des 19. Jahrhunderts ist jedoch bekannt, dass all diese Erscheinungen keine Anhaltspunkte für die Unterscheidung essbarer und giftiger Pilze bieten.
Bis in die Neuzeit hinein wurde das Erscheinen von Pilzen mit Miasmen erklärt: Pilze entstünden durch schlechte Ausdünstungen der Erde oder durch faulenden Untergrund. Auch der Glaube an die Urzeugung (generatio spontanea) wurde durch Pilze genährt, weil man ihre Sporen vor der Erfindung des Mikroskops nicht sehen konnte. Adamus Lonicerus schrieb, dass bestimmte Pilze Schwämme der Götterkinder seien, weil sie ohne einen Samen wüchsen, daher würden sie auch von den Poeten Gygenais, terra nati (Kinder der Erde), genannt.
Zum lange Zeit eher sinistren Bild der Pilze in der Öffentlichkeit haben früher unerklärliche Phänomene wie der Hexenring und das nächtliche grüne Leuchten des Hallimasch-Myzels beigetragen.
Rekorde

Der größte bekannte Pilz der Welt ist ein Dunkler Hallimasch. Er befindet sich in einem Naturschutzgebiet in Oregon und wird mit einer Ausdehnung des Myzels über fast Tausend Hektar Wald als das größte bekannte Lebewesen betrachtet. Sein Gewicht wird auf 600 Tonnen geschätzt, sein Alter auf fast 2000 Jahre.
Der Pilz mit dem größten bekannten Fruchtkörper ist ein Feuerschwamm der Art Phellinus ellipsoideus, der im Jahr 2010 in der chinesischen Provinz Hainan gefunden wurde. Der Fruchtkörper war 10,85 Meter lang, 82 bis 88 Zentimeter breit und 4,6 bis 5,5 Zentimeter dick. Untersuchungen der Dichte des Pilzes ergaben, dass der gesamte Fruchtkörper 400 bis 500 Kilogramm wog. Sein Alter wurde auf etwa 20 Jahre geschätzt.
Literatur
- Robert Hofrichter: Das geheimnisvolle Leben der Pilze: Die faszinierenden Wunder einer verborgenen Welt. 3. Auflage. Gütersloher Verlagshaus 2017, ISBN 978-3-579-08676-7.
- Heinrich Holzer: Fadenwesen: Fabelhafte Pilzwelt. Edition Lichtland, 2011, ISBN 978-3-942509-11-4.
- Hans und Erika Kothe: Pilzgeschichten: Wissenswertes aus der Mykologie. Springer, Berlin/Heidelberg 2013, ISBN 978-3-540-61107-3.
- R. Flammer, E. Horak: Pilzvergiftungen. Schwabe Verlag, Basel 2003, ISBN 3-7965-2008-1.
- Christian Rätsch: Pilze und Menschen: Gebrauch, Wirkung und Bedeutung der Pilze in der Kultur. AT Verlag, Aarau (Schweiz) 2010, ISBN 978-3-03800-542-1.
- H. Dörfelt, H. Heklau: Die Geschichte der Mykologie. Einhorn-Verlag, Schwäbisch Gmünd 1998, ISBN 3-927654-44-2.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 12.06. 2026