Blutkreislauf
Der Blutkreislauf, auch Kreislauf, Blutzirkulation und kurz Zirkulation, Blutgefäßsystem oder Blutbahn genannt, ist das Strömungssystem des Blutes im Körper des Menschen und der meisten Tiere, ausgehend vom Herzen zu den Organen und wieder zurück zum Herzen. Herz und Blutkreislauf zusammen bilden das Herz-Kreislauf-System (kardiovaskuläres System). Seine Aufgabe ist die Versorgung der Organe mit Nährstoffen, Signalstoffen und anderem, die Entsorgung von Stoffwechselprodukten und bei den meisten Gruppen mit Kreislauf auch die Versorgung mit Sauerstoff und der Abtransport von Kohlendioxid. Je nach Tiergruppe können weitere Aufgaben hinzu kommen.
Blutgefäße, die Blut zum Herzen leiten, werden als Venen (Blutadern) bezeichnet; diejenigen Gefäße, die Blut vom Herzen zu den Organen leiten, nennt man Arterien (Schlagadern). Diese Bezeichnungen gelten unabhängig davon, ob das Blut im jeweiligen Gefäß sauerstoffarm oder sauerstoffreich ist. Je stärker sich die Blutgefäße verzweigen, desto kleiner wird ihr Durchmesser. Bei Tieren mit einem geschlossenen Kreislaufsystem werden Arterien zuerst zu Arteriolen und diese zu Kapillaren, in welchen der größte Teil des Stoffaustausches mit den Geweben stattfindet. Diese führen wiederum zusammen und bilden die postkapillären Venolen, die sich zu Venen vereinigen.
Bei Tieren mit einem offenen Kreislaufsystem ergießt sich die Flüssigkeit, die hier nicht Blut, sondern Hämolymphe genannt wird, aus arteriellen Blutgefäßen in die Körperhöhle, um die Organe zu umfließen. Durch die Körperhöhle fließt sie zu venösen Gefäßen oder direkt zurück zum Herzen.
Funktion
Stofftransport
Bei etlichen Gruppen der vielzelligen Tiere sichert ein Kreislauf das Überleben des Organismus, indem er Stoffwechsel in allen Teilen des Körpers ermöglicht und die chemischen und physiologischen Eigenschaften der Körperflüssigkeiten aufrechterhält.
Bei den meisten Tieren mit Kreislauf dient er zum Transport der Atemgase Sauerstoff und Kohlenstoffdioxid (Ausnahme: Insekten). Dann transportiert er sauerstoffreiches Blut oder Hämolymphe aus den Atmungsorganen (Lungen, Kiemen oder Haut) in die Gewebe und sauerstoffarme, kohlendioxidreiche Flüssigkeit zurück zu den Atmungsorganen (siehe auch Atmung). Zur Unterstützung dieser Prozesse können Sauerstofftransporter vorhanden sein; bei den Wirbeltieren ist dies Hämoglobin, das in den roten Blutzellen verpackt ist.
Aus der Verdauung gewonnene Nährstoffe wie Fette, Zucker oder Eiweiße werden aus dem Verdauungstrakt in die einzelnen Gewebe transportiert, um dort je nach Bedarf verbraucht, weiterverarbeitet oder gespeichert zu werden. Die entstandenen Stoffwechsel- oder Abfallprodukte (zum Beispiel Harnstoff oder Harnsäure) werden in andere Gewebe oder zu den Ausscheidungsorganen (bei Wirbeltieren Niere und Leber) transportiert.
Außerdem kann ein Transport von Botenstoffen wie Hormonen, Zellen der Körperabwehr und Bestandteilen des Gerinnungssystems stattfinden. Über die gleichmäßige Verteilung des Blutes oder der Hämolymphe durch den Körper wird auch der pH-Wert, die extrazelluläre Ionenkonzentration und die Osmolarität der Körperflüssigkeiten reguliert (Homöostase).
Andere Funktionen
In manchen Tiergruppen werden vom Kreislaufsystem neben dem Stofftransport noch weitere Funktionen erfüllt. Hierzu gehören eine Temperaturregulation innerhalb des Körpers (Thermoregulation) und sehr verschiedene hydraulische Funktionen. Beispiele sind das Strecken der Beine bei Spinnen durch hydrostatischen Druck[1] und die Erektion der Geschlechtsorgane bei Wirbeltieren durch die erhöhte Blutmenge in Schwellkörpern sowie die Bereitstellung des Drucks für die Filtration des Harns in der Niere.[2]
Formen von Kreislaufsystemen
Diffusion ist über längere Entfernungen sehr langsam, da die Dauer mit dem Quadrat der Entfernung zunimmt. Beispielsweise braucht ein Glucosemolekül über acht Minuten, um bei 37 °C in Wasser über einen Millimeter zu diffundieren. Daher haben alle vielzelligen Tiere, die mehr als nur wenige Zellschichten dick sind, Möglichkeiten entwickelt, einen Flüssigkeitsstrom zu erzeugen,[3] der einen Transport per Konvektion erlaubt.
Tiere ohne Blutkreislauf
_anterior.jpg)
Die Tiergruppen der Schwämme, Nesseltiere, Fadenwürmer und Plattwürmer können auch ohne Kreislaufsystem einen Flüssigkeitsstrom kreieren. Schwämme erzeugen mit Hilfe von sich bewegenden Flagellen einen Strom durch die Körperhöhle. Nesseltieren gelingt dies durch Muskelkontraktionen. Plattwürmer setzen ihre Gewebsflüssigkeit mit der Hilfe von Zilien in Bewegung. Bei den Nesseltieren und Plattwürmern kommen weitverzweigte Darmsysteme vor, die als Gastrovascularsysteme („Magengefäßsystem“) bezeichnet werden, und die das Fehlen eines Blutgefäßsystems zum Teil ausgleichen können.[4] Fadenwürmer setzen ihre innere Flüssigkeit durch Muskelkontraktionen in Bewegung.[3]
Zu den Tieren, die keinen Kreislauf haben, gehören auch einige Vielborster und Egel (beides Gruppen der Ringelwürmer), viele Ruderfußkrebse, Rankenfußkrebse und Pfeilwürmer, die alle die Flüssigkeit in der sekundären Leibeshöhle (Coelom) wie die Fadenwürmer durch Kontraktion der Körperwandmuskulatur in Bewegung bringen. Hüpferlinge dagegen nutzen hierfür die Bewegung der Gliedmaßen, der Vielborster Tomopteris macht dies durch Flimmerung. Man spricht in diesen Fällen von einem Coelomkreislauf.[5]
Unterscheidung von offenen und geschlossenen Kreislaufsystemen
Bei anderen mehrzelligen Tiergruppen gibt es zwei Typen von Kreislaufsystemen. Bei geschlossenen Kreislaufsystemen fließt Blut vom Herzen über Arterien, Kapillaren und Venen zurück zum Herzen in Blutgefäßen. Bei offenen Kreislaufsystemen verlässt die Flüssigkeit, die dann nicht Blut, sondern Hämolymphe genannt wird, die Gefäße, um über unterschiedlich lange Strecken durch Lückensysteme zwischen den Geweben zurück zum Herzen zu fließen.[5]
Die genaue Abgrenzung ist in der Literatur uneinheitlich. In einem Lehrbuch von 2009[5] findet sich die Aussage, dass bei geschlossenen Kreislaufsystemen das Blut ausschließlich durch mit Endothel ausgekleidete Blutgefäße fließe. Als Beispiele für einen geschlossenen Kreislauf werden unter anderen Schnurwürmer, einige Ringelwürmer und alle Wirbeltiere erwähnt. Bei den Kopffüßern sei der Kreislauf „nahezu“ geschlossen. In einer Übersichtsarbeit von 2012[2] heißt es dagegen, dass echtes Endothel nur bei Wirbeltieren vorkomme und die Gefäße der Wirbellosen von extrazellulärer Matrix ausgekleidet seien. Zwar würden auch hier bei manchen Arten Zellen innerhalb der Basalmembran vorkommen, die Auskleidung sei jedoch unvollständig und diese Zellen hätten keine Verbindungskanäle (Gap Junctions) zueinander, es sei demnach kein echtes Endothel vorhanden.
Auch in dieser Arbeit wird allen Wirbeltieren ein geschlossenes Kreislaufsystem zugesprochen, aber auch einigen Wirbellosen, namentlich Ringelwürmern und Kopffüßern. Es wird ausdrücklich erwähnt, dass die Abgrenzung zwischen offenen und geschlossenen Kreislaufsystemen nicht immer eindeutig und eine zu starke Vereinfachung sei. Denn bei manchen Tieren mit offenem Kreislauf wie dem Hummer (siehe unten) oder der marinen Seeohren (Haliotis), bei der die Hinterleibsarterien in kapillar-ähnliche Gefäße verzweigen, gäbe es durchaus auch Merkmale eines geschlossenen Kreislaufes. Und auch bei geschlossenen Kreisläufen der Wirbeltiere gäbe es Gefäßbereiche, wo es direkten Kontakt des Blutes mit dem Interstitium gäbe (siehe auch unten). Ein weiteres Lehrbuch von 2008[6] führt wiederum aus, dass eine Gruppe der Wirbeltiere, nämlich die Kieferlosen (Schleimaale und Neunaugen), im Gegensatz zu den anderen Wirbeltieren ein teilweise offenes Kreislaufsystem haben, bei dem sich das Blut in einigen Körperbereichen in offene Lakunen entleert. Den höheren Kopffüßern (Kalmare und Kraken) wird ein geschlossenes Kreislaufsystem zugesprochen.
Offene Kreislaufsysteme
Hämolymphe als einheitliche Körperflüssigkeit
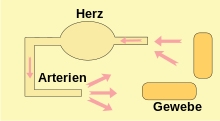
Bei verschiedenen Gruppen wirbelloser Tiere findet man ein offenes Kreislaufsystem, bei dem die Hämolymphe vom Herzen in mehr oder weniger kurze Arterien und von dort in die Körperhöhlen gepumpt wird. Kapillaren fehlen meist, stattdessen umströmt die Hämolymphe die Gewebe, bis es schließlich in offene Venen und durch diese zum Herzen zurück fließt oder direkt aus der Körperhöhle ins Herz zurückkehrt. Die Hämolymphe fließt dabei langsam und mit geringem Druck. Das bewegte Flüssigkeitsvolumen ist relativ groß, da die extrazelluläre Flüssigkeit fast vollständig am Kreislauf beteiligt ist. Bei Schnecken entspricht es etwa 50 % des Körpervolumens.[7]
Bei der als Kalifornischer Seehase (Aplysia californica) bekannten Schneckenart liegt der Anteil der Hämolymphe am Körpergewicht sogar bei 79,3 %, bei der Gemeinen Strandkrabbe (Carcinus maenas) bei 37 % und bei der Amerikanischen Großschabe (Periplaneta americana) bei 19,5 %. Generell macht sie bei Insekten 15–40 % des Gewichts aus. Dabei ist zu berücksichtigen, dass diese Flüssigkeitsmenge die gesamte extrazelluläre Flüssigkeit beinhaltet. Bei Tieren mit geschlossenem Kreislauf gehört zur Vergleichsmenge daher nicht nur das Blutvolumen, sondern auch Gewebsflüssigkeit und die Lymphe.[8]
Konsequenzen und Vorteile eines offenen Kreislaufsystems
Die langsame Zirkulation der Hämolymphe ist vermutlich ein Grund dafür, warum die Körpergröße von Tieren mit offenem Kreislauf im Vergleich zu Wirbeltieren beschränkt ist.[8]
Ein geschlossenes Kreislaufsystem erfordert für die Durchblutung einen höheren Blutdruck und somit einen stärkeren Herzmuskel als ein offenes System, bei dem für die langsamere Durchblutung mit niedrigerem Druck ein dünnwandiges Herz genügt.[9]
Die Herztätigkeit hat eine nur geringe Auswirkung auf den sehr variablen Druck. Stattdessen hängt dieser von der Bewegung anderer Muskeln, der Körperhaltung und der Position der inneren Organe ab. Er kann beispielsweise durch Füllung des Darms mit Luft oder Wasser gesteigert werden, was bei Häutungen und bei der Entfaltung von Flügeln oder anderen Körperteilen nach dem Schlüpfen der erwachsenen Tiere (Imago) aus der Puppe oder dem letzten Larvenstadium eine Rolle spielen kann. Bei den Insekten haben außerdem die Atembewegungen des Hinterleibs (Abdomen) eine Wirkung auf den Druck.[10]
Verbreitung
Offene Kreislaufsysteme kommen vor bei Schnecken, Muscheln, Gliederfüßern, Manteltieren und manchen Ringelwürmern.
Geschlossene Kreislaufsysteme
Geschlossen bedeutet nicht abgeschlossen: Stoffaustausch
Wie erwähnt wird als geschlossenes Kreislaufsystem ein System bezeichnet, bei dem das Blut vom Herzen zu den Organen und zurück durch Blutgefäße fließt.[5] „Geschlossen“ in diesem Sinne bedeutet nicht, dass das Blut im Gefäßsystem vom restlichen Körper hermetisch abgeschlossen ist. Ein Stoffaustausch findet besonders in den Kapillaren statt, aber nicht nur dort.
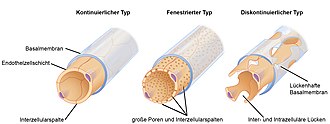
Beispielhaft sollen einige der Stellen erwähnt werden, bei denen bei Wirbeltieren intensiver Stoffaustausch oder Stoffübertritt stattfindet. In deren Leber, Knochenmark und Lymphknoten kommen Kapillaren mit diskontinuierlichem Endothel (Sinusoide) vor.[11] Öffnungen mit bis zu einem Mikrometer Größe zwischen den Endothelzellen der Gefäßwand erleichtern den Stoffaustausch und den Übertritt von Blutzellen.[12] In den Lebersinusoiden bestehen große Lücken in der Auskleidung.[13][14] So können sinusoidale und extravasale Volumina in der Leber des Hundes differenziert werden.[15] Auch zum Lymphgefäßsystem gibt es bei den Wirbeltieren einen oder mehrere offene Übergänge.[16] In der hämochorialen Plazenta (z. B. bei Primaten) münden die mütterlichen Spiralarterien offen und das Blut wird in ein Plazentalabyrinth freigesetzt, wo es die Chorionzotten umspült und von den Spiraladern entwässert wird.
Vorteile eines geschlossenen Kreislaufsystems
Trotz dieser Übergänge zu anderen Systemen wird ein solches Kreislaufsystem als geschlossen bezeichnet,[17] denn das Blut fließt im Gegensatz zu offenen Kreislaufsystemen nicht durch die Körperhöhle oder Spalten zwischen den Geweben zurück zum Herzen und durchmischt sich auch nicht vollständig mit der Gewebsflüssigkeit wie bei offenen Kreislaufsystemen.
Durch die Trennung von Blut und Gewebsflüssigkeit ist das zu transportierende Flüssigkeitsvolumen vergleichsweise klein. Beispielsweise entspricht es bei Tintenfischen etwa 15 % des Körpervolumens, beim Menschen nur 6–8 %.[7] Beim Riesenerdwurm Glossoscolex giganteus (ein Ringelwurm) beträgt es 6,1 %, beim Kraken Octopus honkongensis 5,8 % und beim Teichfrosch (Rana esculenta) 5,6 %.[8]
Geschlossene Kreislaufsysteme haben sich in mehreren Tierstämmen unabhängig voneinander entwickelt. Meist kommen sie bei sehr aktiven oder bei in sauerstoffarmen Umgebungen lebenden Tieren vor. Die Entwicklung erfolgte parallel zu Sauerstofftransportern im Blut. Dadurch müssen nur geringe Blutmengen bewegt werden, um den Sauerstoffbedarf zu stillen.[18]
Wichtige Vorteile geschlossener Systeme für Tiere mit aktivem Lebensstil sind die effektive Verteilung des Bluts in alle Körperregionen und die Möglichkeit zur gezielten Regulation der Durchblutungssteuerung der Organe, um besonders hohen Bedarf an einer Stelle decken zu können, ohne die Gesamtmenge des bewegten Blutes unnötig zu erhöhen. Auch eine parallele Durchblutung der Organe (statt serieller Durchblutung) und somit Durchblutung aller Organe mit sauerstoffreichem Blut ist nur mit einem geschlossenen System möglich.[2]
Verbreitung
Geschlossene Kreislaufsysteme kommen vor bei Ringelwürmern, Seegurken, Schädellosen, Wirbeltieren und Kopffüßern, aber nicht notwendigerweise bei allen Vertretern der Gruppe.
Weichtiere (Mollusca)
Bei den Weichtiere haben Schnecken, Muscheln und der urtümliche Kopffüßer Nautilus (ein Perlboot) ein offenes Kreislaufsystem. Die meisten Kopffüßer haben jedoch ein geschlossenes.[19]
Schnecken und Muscheln
Muscheln und Schnecken haben ein kräftiges Herz, das meist aus zwei Abschnitten besteht, Vorhof und Kammer. Der erzeugbare Druck und damit die Flussgeschwindigkeit ist für offene Kreislaufsysteme vergleichsweise hoch. Bei der Weinbergschnecke kann er während der Kontraktion 19 mmHg (2,5 kPa) erreichen. Bei ihr wird das Blut über lange Gefäße transportiert, bevor es in die Körperhöhle (Hämocoel) austritt. Manche Muscheln nutzen den hohen Druck zur Bewegung des Fußes und damit zur Fortbewegung. Mit dem Blut werden auch die Atemgase transportiert.[9]
Kopffüßer
Die meisten Kopffüßer, nämlich Kalmare, Kraken und Sepien, haben ein geschlossenes Kreislaufsystem. Dieses hat sich vermutlich aus einem offenen entwickelt, wie es bei Nautilus vorkommt. Der geschlossene Kreislauf der Kalmare und Kraken hat drei gekammerte Herzen. Vom Körperherz fließt sauerstoffreiches Blut in die Organe, von dort das sauerstoffarme Blut zu den beiden Kiemenherzen, die es zu den Kiemen leiten. Von dort kehrt das Blut zum Körperherz zurück.[19]
Bei manchen Arten wurden zusätzliche peristaltische Kontraktionen in den Gefäßen der Kiemen, in den Armen oder in der Hohlvene beobachtet. Entsprechend ihrer beweglicheren Lebensweise haben Kopffüßer deutlich höhere Drücke im Kreislauf als Schnecken und diese wiederum höhere als Muscheln. Beim Kraken Octopus dofleini wurde in der Aorta ein Blutdruck von 45 zu 30 mmHg (6 und 4 kPa) gemessen. Bei dem Gewöhnlichen Kraken (Octopus vulgaris) wurde gezeigt, dass die Kiemenherzen synchronisiert jeweils kurz nach dem Körperherz schlagen. Außerdem wurde beobachtet, dass Atembewegungen den Kreislauf zusätzlich antreiben.[20]
Ringelwürmer (Annelida)
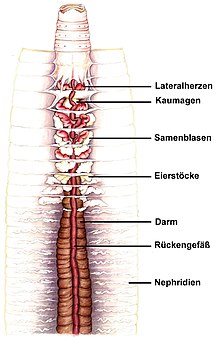
Bei den Ringelwürmern kommen sowohl offene als auch geschlossene Kreislaufsysteme vor. In beiden enthält die Flüssigkeit Sauerstofftransporter.
Regenwürmer und andere Wenigborster
Wenigborster wie der Regenwurm haben einen geschlossenen Kreislauf mit einem Haupt-Bauchgefäß und einem Haupt-Rückengefäß, die beide längs des Tiers verlaufen. Sie sind durch kleinere Gefäße verbunden. Das Rückengefäß kann sich rhythmisch-peristaltisch zusammen ziehen und dadurch das Blut Richtung Vorderende bewegen. Dort sind mehrere Ringgefäße mit Lateralherzen, die eine Strömung zum Bauchgefäß erzeugen, in welchem ein Fluss zum Hinterende herrscht. Durch die erwähnten kleineren verbindenden Gefäße kehrt das Blut zurück in das Rückengefäß.[19]
Es gibt demnach als Besonderheit bei den Ringelwürmern kein zentrales Herz, sondern mehrere kontrahierende Abschnitte im Blutgefäßsystem. Gasaustausch mit der Luft findet über die Haut statt. Sauerstoffarmes Blut wird durch kleinere Gefäße vom Bauchgefäß zur gut durchbluteten Haut geleitet und von dort weiter zum Rückengefäß.[9]
Gut untersucht ist der Riesenerdwurm Glossoscolex giganteus, der bei einem Durchmesser von 2–3 cm eine Länge von 1,2 Metern und ein Gewicht von 0,5 bis 0,6 Kilogramm erreicht. Die peristaltischen Kontraktionen des Rückengefäßes können einen Druck von 10–18 mmHg (1,4–2,4 kPa) aufbauen. Die in der Regel synchron schlagenden 5 Paare der Lateralherzen können im Bauchgefäß einen Druck von 80 mmHg oder über 10 kPa aufbauen, ein Wert der dem arteriellen Blutdruck der Wirbeltiere nahe kommt. Dieser hohe Druck dient zur Durchblutung des Hautmuskelschlauches und der Metanephridien. Der Blutdruck ändert sich während der Bewegung des Tiers sehr stark. Die höchsten Werte werden erreicht, wenn es sich streckt, um während der Verkürzung des Tieres auf etwa halbe Werte zu sinken.[9]
Die Anzahl der Kontraktionswellen im Rückengefäß von Glossoscolex giganteus lag in Ruhe bei etwa acht pro Minute, beim Gemeinen Regenwurm (Lumbricus terrestris) bei etwa 11 pro Minute. Die Rate wird erhöht bei körperlicher Anstrengung und beeinflusst von der Füllmenge des Gefäßes. Die Füllmenge beeinflusst auch die Kraftentwicklung der Kontraktion. In elektronenmikroskopischen Bildern von Blutgefäßen des gemeinen Regenwurms sieht man diese angefüllt mit Hämoglobin-Partikeln, die frei im Blut schwimmen.[2]
Vielborster
Vielborster, meist im Meer lebende Tiere, haben ein ähnliches Kreislaufsystem wie die Wenigborster, jedoch haben die Blutgefäße oft eine dünnere Wand. Nicht immer sind die Gefäße mit Endothel (oder je nach Definition mit endothel-ähnlichen Zellen) ausgekleidet, daher ist es nach einer möglichen Definition ein offenes Kreislaufsystem. Bei manchen Arten kommen zusätzliche Nebenherzen in der Peripherie des Tieres vor, besonders an den Kiemen.[9]
Egel
Bei der dritten Gruppe der Ringelwürmer, den Egeln, kommt nur bei den Rüsselegeln ein echtes Blutgefäßsystem vor. Bei den anderen Egeln ist es reduziert, die Transport- und Versorgungsaufgaben werden von einem geschlossenen Coelomsystem übernommen. Dessen Seitengefäße und Bauchgefäß können sich rhythmisch zusammenziehen. In der Flüssigkeit ist Hämoglobin gelöst. Beim gut untersuchten Blutegel (Hirudo medicinalis) wirken die Seitengefäße als sich peristaltisch kontrahierende Herzen, die einen Blutstrom Richtung Vorderende und einen Druck bis zu 100 mmHg erzeugen. Klappen und Sphinkter an den Enden verhindern einen Rückfluss. In Rücken- und Bauchgefäß herrscht Fluss Richtung Hinterende.[21]
Schnurwürmer (Nemertea oder Nemertini)
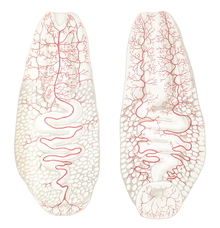
Bei den Schnurwürmern, einer Gruppe im Meer lebender Arten, kommt ein Gefäßsystem vor, das zwar oft als Blutgefäßsystem bezeichnet wird, jedoch wahrscheinlich eine Abwandlung des Coeloms darstellt, also eine Bildung der sekundären Leibeshöhle. Es ist somit nicht dem primären Blutgefäßsystem der Bilateria homolog. Die Hauptgefäße liegen seitlich der Länge nach im Tier, die am Vorderende und in der Nähe des Afters bauchseits verbunden sind. Manche Arten haben außerdem ein Rückengefäß oder eine Aufspaltung der Hauptgefäße. Die größeren Gefäße können sich zusammen ziehen, zur Flüssigkeitsbewegung trägt auch die Körpermuskulatur bei.[22]
Gliederfüßer (Arthropoda)
Gliederfüßer, zu denen Insekten, Tausendfüßer, Krebstiere und Spinnentiere gehören, haben alle ein offenes Kreislaufsystem.[23]
Krebstiere (Crustacea)
_(18221234840).jpg)
Bei den Krebstieren reicht die Spanne der Kreislaufsysteme von sehr einfach bis sehr komplex. Die wahre Beschaffenheit der Blutzirkulation bei den Krebstieren (und bei den Insekten) erkannte der Mediziner und Naturforscher John Hunter.[24] Der Kiemenfußkrebs Branchinecta hat ein röhrenförmiges Herz, welches fast den ganzen Körper durchzieht, und vergleichsweise wenige Gefäße.
Zehnfußkrebse
Eine Besonderheit kommt bei den Zehnfußkrebsen vor. Diese haben zwar ebenfalls anatomisch gesehen ein offenes Kreislaufsystem, jedoch funktionell viele Aspekte eines geschlossenen Systems: kleine Blutgefäße entsprechen Kapillaren und die Hämolymphe fließt durch definierte Kanäle durch das Gewebe zum Herz zurück, die bei einigen Arten so klein sind, dass sie Blutgefäßen funktionell entsprechen. Zahlreiche Blutgefäße haben muskulöse Ventile, mit deren Hilfe die Durchblutung der nachgeschalteten Organe reguliert wird. Das Herz ist eine muskulöse, kontraktile Kammer und liegt in einem Sack, dem Perikardialsinus oder Perikard. Die Hämolymphe fließt über mehrere Arterien von dort in zahlreiche Körperregionen, um schließlich tief im Gewebe aus den Gefäßen auszutreten. Ein Sinus auf der Bauchseite sammelt die Hämolymphe und führt sie zu den Kiemen. Nach Sauerstoffaufnahme gelangt sie über Venen zum Perikardialsinus, wo sie durch kleine Öffnungen, die Ostien, zurück ins Herz gelangt.[3][23]
Das Herz dieser Krebse kann Drücke von 8–15 mmHg (1–2 kPa), beim Hummer bis zu 20 mmHg,[2] erzeugen und auch beim Erschlaffen des Herzmuskels sinkt der Druck in der Aorta nicht auf null, sondern bleibt durch einen Windkesseleffekt der großen Gefäße bei etwa 1,5 mmHg (0,2 kPa) und damit höher als in der offenen Körperhöhle (Hämocoel), so dass ein kontinuierlicher Flüssigkeitsstrom gewährleistet wird. Dadurch erreicht ein Hummer ein Herzminutenvolumen von 10–30 ml pro Minute und eine Umlaufzeit seiner Hämolymphe von 2–8 Minuten.[9]
Der Herzschlag wird von Nervenzellen gesteuert (neurogenes Herz), und nicht durch spezialisierte Muskelzellen wie bei den Wirbeltieren. Das Herzganglion liegt rückenwärts am Herzen auf und wird seinerseits vom Zentralnervensystem gesteuert.[9]
Bei Hummern wurden Herzfrequenzen zwischen 61 und 83 Schlägen pro Minute beobachtet. In seiner Hämolymphe gelöst ist der kupferhaltige Sauerstofftransporter Hämocyanin und es wurden drei bis elf morphologisch unterscheidbare Haemocyten gefunden, deren Aufgaben vermutlich in der angeborenen Immunantwort und der Blutstillung liegen.[2]
Andere Krebstiere
Innerhalb der Krebstiere wurde außer bei den Zehnfußkrebsen Fluss in Gefäßen über weite Strecken nur bei den Asseln beobachtet. Bei den Daphnien ähnelt das Herz dem der Zehnfußkrebse, es sind jedoch keine Gefäße vorhanden. Das Röhrenherz der Kiemenfüßer, zu denen Artemia gehört, ähnelt eher dem der Insekten. Bei den Rankenfußkrebsen, darunter Seepocken und Entenmuscheln, gibt es kein Herz im eigentlichen Sinn. Die Muskulatur des Thorax übernimmt die Pumpfunktion.[9]
Insekten (Insecta)

Insekten haben trotz einer hohen Stoffwechselrate ein eher einfaches, offenes, in seiner Beschaffenheit von dem Mediziner und Naturforscher John Hunter entdecktes[25] Kreislaufsystem. Dies ist möglich, weil Insekten im Gegensatz zu den meisten Tieren mit Kreislauf diesen nicht zum Atemgastransport nutzen, sondern nur zum Transport von Nährstoffen, Immunzellen, Signalmolekülen und dergleichen. Der Gasaustausch geschieht über ein Tracheensystem, hohle Röhren, die von der Körperoberfläche direkt an die Gewebe heranführen.[23]
Bei vielen Insekten besteht das Gefäßsystem nur aus einem entlang dem Rücken liegenden langen Herzen, das hinten verschlossen ist und nach vorne in die Aorta übergeht. Kontraktionswellen treiben die Hämolymphe Richtung Kopf, wo sie in einen Sinus austritt und zu einem weiteren Sinus sickert, um durch diesen zurück in den Körper zu fließen. Körperbewegungen halten den Fluss in Gang, so dass wie bei anderen Arthropoden ein Übergang über Ostien zurück ins Herz erfolgt. Wenn das Herz erschlafft, wird es durch elastische Fasern des Aufhängeapparats gedehnt, so dass die Hämolymphe durch Unterdruck einströmt. Bei der Kontraktion werden die Ostien durch Klappen aus Bindegewebe verschlossen. Bei der Wanderheuschrecke Locusta migratoria wurde ein maximaler Druck von nur etwa 7 mmHg (0,9 kPa) gemessen.[23][9]
Häufig kommen in den Antennen, Flügeln und Beinen zusätzliche, einfache Herzen vor, bei manchen Arten können es mehrere Dutzend sein. Sie dienen der Versorgung von besonders stoffwechselaktiven Geweben und werden auch Nebenherzen oder pulsatile Organe genannt.[23][9]
Um auch die Enden der langen, dünnen Gliedmaßen durchfließen zu können, sind diese der Länge nach durch Septen in zwei Bereiche unterteilt. Nur am Ende der Extremität findet sich eine Öffnung, so dass die Hämolymphe auf der einen Seite hinein und auf der anderen wieder zurück strömt.[9]
Bei einigen Arten der Schmetterlinge, bei der Schmeißfliege Calliphora, den Goliathkäfern und dem Nashornkäfer wurde beobachtet, dass sich die Schlagrichtung des Herzens umkehren kann. Das Herz hat bei diesen Arten hinten eine oder zwei Öffnungen, durch die die Hämolymphe dann austreten kann. Meist wird häufiger Blut nach vorne gefördert als nach hinten. Durch die abwechselnde Richtung kann die Hämolymphe zwischen Vorder- und Hinterteil pendeln, was auch zu einer verstärkten Belüftung der Tracheen führt.[10]
Im Gegensatz zu allen anderen wirbellosen Tieren haben Insekten eine effektive Blut-Hirn-Schranke entwickelt. Die Diffusion von wasserlöslichen Molekülen von der Hämolymphe in die unmittelbare Umgebung der Nervenzellen ist dadurch stark eingeschränkt.[10]
Spinnentiere (Arachnida)
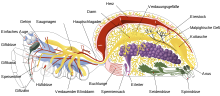
Im Gegensatz zu den Insekten müssen Spinnentiere (darunter Spinnen, Milben, Skorpione) wie die Krebstiere nicht nur Nährstoffe, sondern auch Atemgase mit der Hämolymphe transportieren. Das im Hinterleib (Opisthosoma) am Rücken liegende Röhrenherz ist dem der Insekten ähnlich. Gefäße erlauben eine gezielte Durchblutung der Buchlunge. Eine Besonderheit liegt bei Spinnen vor, da sie ihre Beine nicht mit Muskelkraft strecken, sondern hydraulisch über einen relativ hohen Druck der Hämolymphe, die über eigene Blutgefäße dorthin gelangt.[9][1]
Stachelhäuter (Echinodermata)
_(20336762836)-Fig7.jpg)

Stachelhäuter sind die einzigen ursprünglich bilateralen Tiere, die eine Fünfersymmetrie entwickelt haben. Die Larven sind jedoch bilateral. Alle zugehörigen Gruppen leben ausschließlich im Meer, darunter Seesterne, Seeigel, Schlangensterne und Seegurken. Die Stachelhäuter haben zwei Gefäßsysteme. Ein wesentliches Merkmal ist ihr Ambulacralsystem, manchmal auch als Wassergefäßsystem bezeichnet. Es enthält jedoch kein Meerwasser, sondern eine Körperflüssigkeit. Es hat einen ringförmigen Zentralkanal, von dem je ein Radiärkanal in die fünf Körperstrahlen abgeht. Dieses Gefäßsystem ist eine Neubildung der Stachelhäuter und entspricht nicht dem Blutgefäßsystem der anderen Tiergruppen.[26]
Daneben gibt es das Blutgefäßsystem, das bei den Stachelhäutern auch als Blutlakunensystem oder Haemallakunensystem bezeichnet wird. Endothel kommt nicht vor, manche Gefäßwände enthalten Myofilamente. Die Gefäße enthalten eine hohe Konzentration von Glykoproteinpartikeln und Blutzellen, sogenannte Amöbocyten und Coelomocyten. Der größere Anteil dieser Zellen befindet sich jedoch in Coelomräumen und im Bindegewebe. Einige Blutzellen sind phagozytierend, andere transportieren Sauerstoff. Bei den Seegurken kommen scheibenartige, hämoglobinhaltige Zellen vor. Unklar ist, ob, wie stark, in welche Richtung und wie regelmäßig das Blut in den Gefäßen fließt. Mindestens teilweise fließt es wohl gar nicht oder wenig.[27]
Wesentliche Teile des Blutgefäßsystems sind ein „oraler Gefäßring“, der um den Mund herum läuft (im Tier unten oder vorne) und ein „aboraler Gefäßring“, die Entsprechung am anderen Körperende (siehe Abbildung). Beide sind durch das Axialorgan verbunden, eine Struktur, die viele Blutgefäße und Blutzellen enthält und durch letztere häufig dunkel gefärbt ist. Ihm angelagert ist die Dorsalblase, die pulsiert und vermutlich die Funktion eines Herzens übernimmt. Sie ist auch mit den Gefäßen des Darms und der Gonaden verbunden. Das Axialorgan hat auch Ausscheidungsfunktionen und ist vermutlich den Glomeruli der Wirbeltierniere homolog.[27]
Chordatiere (Chordata)
Urchordaten
Die Manteltiere (Tunicata) und die Schädellosen (Lanzettfischchen), die wie die Wirbeltiere zu den Chordatieren gehören, haben ebenfalls ein offenes Kreislaufsystem. Ein einfaches, röhrenförmiges Herz geht in zahlreiche wohldefinierte Kanäle über, die aber keine Gefäßwände haben, und daher nicht als Blutgefäße angesehen werden. Bei manchen Manteltieren, zum Beispiel Ciona, kann sich die Richtung der peristaltischen Kontraktionen des Herzens umkehren, und somit auch die Flussrichtung. Bei den Lanzettfischchen ist der Kreislauf weitgehend geschlossen, die Gefäße münden nur in wenige Lakunen.[3][23]
Das Lanzettfischchen Branchiostoma lanceolatum hat keine Blutzellen und kein Hämoglobin oder andere Sauerstofftransporter. Ein zentrales Herz gibt es nicht, das Blut wird durch mehrere kontraktile Blutgefäße angetrieben. Am Bauch fließt es nach vorne und am Rücken zurück.[2]
Rundmäuler – kieferlose Fische (Cyclostomata, Agnatha)
Auch eine Gruppe von Wirbeltieren hat ein teilweise offenes Kreislaufsystem, und zwar die Rundmäuler, zu denen die kieferlosen Fische gehören, Neunaugen und Schleimaale. Vom systemischen Herzen kommend verbleibt das Blut in manchen Geweben wie bei anderen Wirbeltieren in Blutgefäßen, aber in anderen Geweben tritt es in offene Blutlakunen über. Die Schleimaale haben in manchen Bereichen des Körpers noch zusätzliche, akzessorische Herzen. Wie bei den anderen Wirbeltieren sind die Blutgefäße mit Endothel ausgekleidet.[23]
Von allen Wirbeltieren haben Schleimaale den niedrigsten arteriellen Blutdruck (6–10 mmHg) und das höchste relative Blutvolumen (18 % der Körpermasse). Schleimaale haben mehrere Sinuse, die mit den Blutgefäßen in Verbindung stehen. Der größte liegt am Rücken und zieht sich von der Schnauze bis zur Schwanzflosse. Er liegt zwischen Skelettmuskulatur und Haut. Das Sinussystem kann bis zu 30 % des gesamten Blutvolumens aufnehmen. Vermutlich kann durch Kontraktion von Skelettmuskeln ein aktiver Übertritt in die Blutgefäße erzeugt werden.[2]
Das Herz der Schleimaale hat einen Vorhof und eine Kammer, aber keine Koronargefäße für die eigene Sauerstoffversorgung. Stattdessen durchströmt das Blut im Herzen Kanäle und Lakunen in der Herzwand. Die Herzschlagfrequenz liegt bei 20–30 Schlägen pro Minute. Das Herzzeitvolumen erreicht mit 8–9 ml pro Minute und Kilogramm Körpergewicht Werte, die sich denen der echten Knochenfischeannähern. Das Herz hat einen Frank-Starling-Mechanismus, wird aber nicht über regulatorische Nerven beeinflusst, wie das bei den anderen Wirbeltieren der Fall ist. Ein zweites gekammertes Herz, das ebenfalls aus Herzmuskelzellen aufgebaut ist, transportiert Blut vom Darm in die Portalvene der Leber. Es schlägt in einem anderen Rhythmus als das systemische Herz.[2]
Das Kreislaufsystem der Neunaugen ähnelt jenem der kiefertragenden Fische: Blut gelangt vom Herzventrikel in die Aorta ventralis von dort in die Kiemenarterien, weiter in die Aorta dorsalis, in der es Richtung Schwanz fließt. Von dieser gehen Arteriolen und schließlich Kapillaren ab, durch Venen geschieht der Rückfluss zum Herzen.[23]
Gemeinsamkeiten der kiefertragenden Wirbeltiere (Gnathostomata)
Kiefertragende Wirbeltiere (Kiefermäuler) besitzen einen geschlossenen Kreislauf. Hier fließt das Blut durch ein kontinuierliches Netz aus Blutgefäßen, das alle Organe erreicht. Die Kapillar-Endothelien der Wirbeltiere können kontinuierlich, fenestriert[28] oder diskontinuierlich sein. Diese Kreisläufe sind in den folgenden Abschnitten beschrieben.
Fische
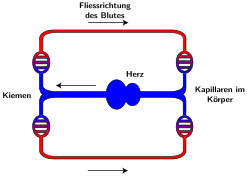
rot = sauerstoffreiches Blut
blau = sauerstoffarmes Blut
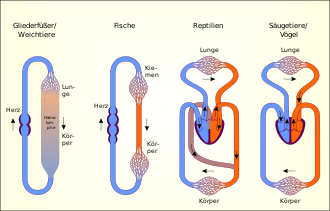
Das Herz-Kreislauf-System der Fische ist unter den kiefertragenden Wirbeltieren das am einfachsten gebaute. Das Herz besteht aus vier Räumen, zwei einleitenden dünnwandigen, Sinus venosus und Vorhof, einer dickwandigen, muskulösen Kammer und dem abschließenden Bulbus oder Conus arteriosus. Zwischen Vorhof und Kammer befindet sich eine Klappe, die einen Rückstrom des Blutes verhindert. Ebenso wie das Herz ist auch der Blutkreislauf selbst relativ einfach strukturiert. Das sauerstoffarme Blut wird aus dem Herzen in die Kiemen gepumpt, in denen es mit Sauerstoff aus dem Wasser angereichert wird. Anschließend wird das sauerstoffreiche Blut in den Körper weiter transportiert. In den Kapillaren gibt es den Sauerstoff ab und nimmt dafür Kohlendioxid auf. Neben dem Herz nimmt auch die Muskulatur der Kiemen am Pumpvorgang teil. Der Nachteil dieser Konstruktion ist, dass der Blutdruck im Kapillarnetz des Kiemenkreislaufs stark abfällt, der Blutstrom durch den Körper also relativ langsam ist. Das Blutvolumen macht weniger als ein Zehntel des Körpergewichts aus. Der Sauerstoffgehalt im Blut der Fische liegt weit unter dem des menschlichen Blutes.
Bei den meisten Fischen sind Herz und Kiemen wie beschrieben in Serie geschaltet. Eine Vermischung von sauerstoffarmen mit sauerstoffreichem Blut findet nicht statt. Der Australische Lungenfisch besitzt wie die Landwirbeltiere einen separaten Lungenkreislauf.[29]
Amphibien
Doppelter Kreislauf
Bei den Amphibien (Lurchen) besteht das Herz aus einer Kammer und zwei Vorhöfen. Der Gasaustausch findet sowohl in der Lunge als auch in der Haut statt. Die beiden Kreisläufe der Amphibien werden daher als Lungen-Haut-Kreislauf und Körperkreislauf bezeichnet. Da sie, im Gegensatz zu Fischen, nicht hintereinander geschaltet sind, spricht man von einem doppelten Kreislauf.
Der linke Vorhof empfängt mit Sauerstoff angereichertes Blut aus der Lunge, der rechte Vorhof eine Mischung von sauerstoffarmem Blut aus dem Körper und sauerstoffreichem Blut aus der Haut. Beide Vorhöfe pumpen das Blut in die einheitliche Kammer. Diese Kammer besitzt einen Ausflusstrakt (Truncus oder Conus arteriosus), der sich in jeweils einen Stamm für die beiden Kreisläufe teilt. Eine leistenartige Erhebung im Ventrikel und im Lumen des Ausflusstrakts sorgt dafür, dass das Blut relativ „sortenrein“ durch das Herz fließt, das Blut aus den beiden Vorhöfen sich also nur wenig vermischt. Das sauerstoffreichere Blut wird zum überwiegenden Teil in die Halsschlagadern und die Aorta gepumpt, während das sauerstoffärmere Blut in die Lungen-Haut-Arterie gelenkt wird. Wie Reptilien und Vögel besitzen die Amphibien bereits einen Nierenpfortaderkreislauf.
Entwicklung
Amphibien haben ursprünglich vier paarige Kiemenbogenarterien, die zu beiden Seiten aus der Aorta entspringen. Bei ausgewachsenen Lurchen entwickelt sich die erste zur Arteria carotis, die den Kopf versorgt. Die Arterien des zweiten Bogens vereinigen sich zur Aorta descendens, der absteigenden Aorta. Die dritte Kiemenbogenarterie bildet sich zurück, und aus den vierten entwickelt sich der paarige Aortenbogen.
Reptilien
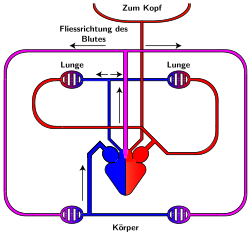
rot = sauerstoffreiches Blut
blau = sauerstoffarmes Blut
rosa = Mischblut
Die meisten zu den Reptilien zusammengefassten Taxa besitzen ein Herz, das wie das der Amphibien aus zwei Vorhöfen und einer Kammer besteht. Diese ist jedoch nahezu vollständig durch eine Scheidewand in zwei Hälften geteilt. Aus dem Körper strömt sauerstoffarmes Blut in den rechten Vorhof, aus den Lungen mit Sauerstoff angereichertes Blut fließt in den linken Vorhof. Beide Vorhöfe pumpen das Blut in die Herzkammer, aus der drei Schlagadern abgehen. In der rechten fließt sauerstoffarmes Blut zur Lunge, in der linken sauerstoffreiches Blut zum Kopf und in den Körper. Da die Trennung der Herzkammer jedoch nicht vollständig ist, kommt es zur Bildung von Mischblut (circa 10 bis 40 Prozent). Dieses fließt durch die mittlere Schlagader in den Körper.
Eine Besonderheit liegt bei den Krokodilen vor, bei denen zwei Herzkammern komplett getrennt sind. Aber zwischen der linken und der rechten Schlagader besteht mit dem Foramen Panizzae ein Verbindungsfenster. Die linke Aorta entspringt an der rechten Herzkammer, die rechte an der linken. Durch das Fenster vermischt sich das sauerstoffreiche Blut der rechten Kammer mit dem sauerstoffarmen der linken Kammer im Bereich der rechten Aorta, so dass Mischblut in den Körperkreislauf (den peripheren Kreislauf[30]) geführt wird und dabei vor allem in die peripheren Bereiche des Körpers gelangt. Zugleich fördert die linke Aorta sauerstoffreiches Blut in den Körper und vor allem in den Kopf des Tieres. Beim Tauchvorgang schließt sich das Foramen Panizzae vollständig, so dass die rechte Aorta nur noch mit sauerstoffarmem Blut versorgt wird, der Kopf jedoch weiterhin sauerstoffreiches Blut bekommt.
Auch bei den Dinosauriern lag vermutlich eine vollständige Trennung der Herzkammern vor. Dies ergibt sich aufgrund ihrer Position im Stammbaum zwischen den Krokodilen und den Vögeln, die beide eine durchgängige Trennwand im Herzen besitzen.
Blutkreislauf der Vögel und Säugetiere
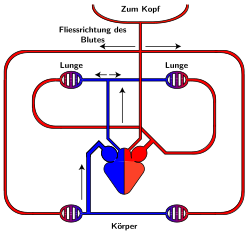
rot = sauerstoffreiches Blut
blau = sauerstoffarmes Blut
Das Herz der Vögel und Säugetiere, also auch das des Menschen, ist vollständig in zwei Hälften geteilt, obwohl es sich um ein Organ handelt. Jede dieser Hälften besteht aus einem Vorhof und einer Kammer, die jeweils als Einheit arbeiten. Insgesamt gibt es also vier Räume. Während die rechte Herzhälfte das Blut durch den (erstmals im 13. Jahrhundert von Ibn an-Nafīs erwähnten[31]) Lungenkreislauf pumpt, der das Blut mit Sauerstoff anreichert, pumpt die linke Herzhälfte das Blut durch den Körperkreislauf, um die Organe mit Nährstoffen und Sauerstoff zu versorgen.
Diese beiden Kreisläufe sind in Reihe geschaltet, so dass das gesamte Blut durch die Lunge fließen muss. Die Organe im Körperkreislauf werden parallel durchströmt.
Ein wichtiger Vorteil eines eigenen, getrennten Lungenkreislaufs ist, dass in diesem ein deutlich geringer Druck herrschen kann als im Körperkreislauf. Bei einem gleich hohen Druck wie im Körperkreislauf würde in der Lunge vermehrt Flüssigkeit aus dem Blut in die Luft übertreten und dadurch den Gasaustausch behindern. Außerdem fungiert die Lunge mit ihren Kapillaren als Filter gegen Blutgerinnsel (Thromben) u.ä., bevor das Blut von der linken Herzseite unter anderem zum Gehirn gepumpt wird. Die Lunge hat dazu thrombenlösende Eigenschaften.
Im Lungenkreislauf verlässt das Blut die rechte Herzkammer über den Lungenstamm (lat. Truncus pulmonalis) in Richtung der Lungen, wo es mit Sauerstoff angereichert wird. Dann wird es von der Lungenvene (lat. Vena pulmonalis) in den linken Herzvorhof gepumpt. Vom linken Vorhof gelangt es in die linke Kammer, von wo aus es durch die Aorta in den Körperkreislauf gelangt. Während bei den Säugern die Aorta auf der linken Körperseite verläuft, liegt sie bei Vögeln auf der rechten. Nach der Versorgung der Organe kehrt das nun mit Kohlenstoffdioxid angereicherte Blut durch die vordere (beim Menschen: obere) und die hintere (beziehungsweise untere) Hohlvene in den rechten Vorhof zurück. Wenn das Blut vom rechten Vorhof in die rechte Kammer kommt, beginnt der Kreislauf von neuem.
Eine Besonderheit stellt das Pfortadersystem der Leber dar. Blut, das von den Organen des Verdauungstrakts kommt, wird in der Pfortader gesammelt und gelangt in die Leber, wo die aufgenommenen Nährstoffe verwertet werden. Das Blut strömt dadurch nacheinander durch zwei Kapillarsysteme, bevor es zum Herzen zurückkommt. Auch die Hirnanhangsdrüse (Hypophyse) hat ein Pfortadersystem. Vögel haben wie Reptilien zudem eine Nierenpfortader.
Entwicklung der Kreislaufsysteme
Phylogenese: Gemeinsamer evolutionärer Ursprung aller Kreislaufsysteme
Kreislaufsysteme finden sich in der Evolution erstmals bei den Bilateria, also Tieren, die zwei klar definierbare mehr oder weniger spiegelbildliche Hälften (rechts und links) haben. Die Bilateria umfassen alle Tiergruppen von Würmern bis zu den Insekten und Wirbeltieren, nicht aber die Schwämme und Nesseltiere.[32]
Der letzte gemeinsame Vorfahr all dieser Gruppen lebte vor etwa 600 bis 700 Millionen Jahren und es wird angenommen, dass es sich um ein segmentiertes, bilaterales Tier handelte. Ein erstes Blutgefäßsystem hat sich womöglich schon zu dieser Zeit entwickelt, möglicherweise um die Trennwände zwischen den Segmenten zu überwinden. Der Fluss wäre durch sich peristaltisch kontrahierende Gefäße in Gang gesetzt worden, vielleicht ähnlich den Rückengefäßen der heutigen Ringelwürmer. Blut wäre vermutlich durch Spalten in der extrazellulären Matrix gesickert, ähnlich einem primitiven geschlossenen Kreislaufsystem. Dieses Modell würde die Homologie aller Kreislaufsysteme im Tierreich bedeuten, die sich alle von diesem ursprünglichen Typus ausgehend entwickelt hätten und demnach einen gemeinsamen phylogenetischen Ursprung hätten. In manchen Tierstämmen hätte sich das Kreislaufsystem vollständig zurück gebildet, etwa bei den Plattwürmern und Fadenwürmern. In anderen Abstammungslinien hätte es sich zu einem offenen Kreislaufsystem entwickelt. Bei einem frühen Kopffüßer wiederum wandelte sich das offene zurück zu einem geschlossenen Kreislaufsystem. Energieverluste bei der Strömung durch Rückfluss und dergleichen führten zu einem Selektionsdruck Richtung der Entwicklung gekammerter Herzen (etwa bei Weichtieren und Wirbeltieren), bei denen diese Problematik vermieden wird.[2]
Der Befund, dass viele Gene und Signalwege eine Rolle bei der Entwicklung des Herz-Kreislaufsystems sowohl bei der Taufliege Drosophila melanogaster als auch bei Wirbeltieren spielen, hat ebenfalls Spekulationen gestützt, dass es einen gemeinsamen Ursprung aller Blutkreislaufsysteme gäbe, die auf einen gemeinsamen bilateralen Vorfahren zurückgehen. Zu den gemeinsamen Signalmolekülen gehören Knochenmorphogenetische Proteine, Fibroblasten-Wachstumsfaktor, Wnt und Notch. Beispielsweise löst der Notch-Signalweg in Hämangioblasten bei Wirbeltieren und bei Drosophila die Bildung von Blutstammzellen aus.[33]
Ontogenese: Kreislaufsysteme werden vom Mesoderm gebildet
Die meisten Coelomata (Tiere mit Coelom) haben ein Gefäßsystem. Dessen Wände entstehen während der Entwicklung des Individuums, der Ontogenese, aus dem Mesothel, bisweilen auch Peritoneum (Bauchfell) genannt. Das Mesothel ist die mesodermale Auskleidung der Körperhöhle (des Coeloms), also eine epitheliale Zellschicht. In Gefäßwänden finden sich kontraktile Myofibrillen, die in manchen Fällen den Blutfluss auslösen. Bei Wirbeltieren und einigen Wirbellosen sind die Blutgefäße mit nicht-muskulärem Endothel ausgekleidet, das von einer Muskelzellschicht umgeben ist. Zwar haben bei den Gliederfüßern und einigen anderen Urmündern (Protostomia) die erwachsenen Tiere kein geschlossenes Coelom, Teile des embryonal angelegten Mesothels überdauern jedoch und bilden epithelial ausgekleidete Gefäßräume. Verglichen mit der oben beschriebenen Situation beim ursprünglichen bilateralen Tier ging ein kontinuierliches Mesothel bei der Entwicklung der Arthropoden in den erwachsenen Tieren verloren. Mesotheliale Zellen bildeten jedoch weiterhin das Röhrenherz am Rücken und die anschließende Aorta.[33]
Bei den Wirbeltieren ist die ontogenetische Entwicklung komplexer. Aus dem embryonalen Mesothel entstehen mehrere Gruppen von Vorläuferzellen, aus denen sich die verschiedenen Gewebeschichten des Herz-Kreislauf-Systems entwickeln, darunter Endothel/Endokard, Myokard und Blutzellen. Diesen Überlegungen zufolge wäre das Drosophila-Herz mit dem Myokard der Wirbeltiere genauso verwandt wie mit dem Endokard und dem Endothel in den Blutgefäßen im Allgemeinen.[33]
Verwandtschaft von Blutgefäßen, Blutzellen und Ausscheidungsorganen
Embryologische und molekulare Hinweise deuten darauf hin, dass Gefäßwandzellen und Blutzellen entwicklungsbiologisch eng miteinander verwandt sind und sich beide aus Hämangioblasten entwickeln. Auch Nephrocyten (Podozyten) sind wohl eng mit den genannten Zelltypen verwandt. Die entsprechenden Systeme für Blutgefäße, Blutzellen und Ausscheidung gehen alle auf das dritte Keimblatt, das Mesoderm zurück, welches bei den frühen Bilateria oder Triploblasten zum ersten Mal auftritt.[33]
Einige heutige Tierstämme der Bilateria haben kein Coelom (Acoelomata, die Plattwürmer) oder kein echtes Coelom (Pseudocoelomata, Fadenwürmer und andere) und kein Gefäßsystem. Auch sie haben jedoch spezielle Ausscheidungsorgane, die Protonephridien. Diese scheinen daher die ältesten epithelialen Gewebe zu sein, die bei den frühen Bilateria aus mesodermalem Parenchym gebildet wurden. Protonephridien bilden ein verzweigtes Röhrensystem, das innen von bewimpertem Epithel und Nephrocyten (hier auch Cyrtocyten genannt) ausgekleidet wird und das entweder nach außen oder in den Verdauungstrakt mündet. Durch den Wimpernschlag wird Flüssigkeit abtransportiert und dadurch ein Unterdruck erzeugt, der weitere Flüssigkeit aus der Körperhöhle ansaugt.[33]
Aufbau der ersten Kreislaufsysteme
Die Coelomata haben in der Regel ein gut ausgebildetes Kreislaufsystem. Die namengebende Körperhöhle, das Coelom, ist meist in mehrere Abschnitte unterteilt (bei Wirbeltieren etwa der Peritonealraum, der Pleuraraum und der Herzbeutel.) Bei einigen zugehörigen wirbellosen Gruppen werden die Gefäße in Spalten zwischen den mesothelialen Trennwänden gebildet, etwa bei Ringelwürmern, Kiemenlochtieren und den Lanzettfischchen. Möglicherweise ist dies die ursprüngliche Bauweise, wie bei den Ringelwürmern, bei denen die beiden Coelom-Räume, die jeweils die linke und rechte Hälfte eines Segmentes um den Darm herum auskleiden, sich am Rücken und Bauch treffen. Auch die Wände der Räume zwischen benachbarten Segmenten treffen sich, zwischen den beiden Mesothelien bleiben jedoch offene Bereiche, die sich verbinden, um Blutgefäße zu bilden. Daraus ergibt sich, dass im Gegensatz zu anderen Röhren oder Öffnungen im Körper, das Innere der primitiven Blutgefäße von der basalen Seite des umgebenden Epithels umgeben wird, und nicht von der apikalen, „äußeren“ Seite. Tatsächlich lässt sich bei den wirbellosen Tiergruppen eine dem Lumen zugewandte, ausgeprägte Basalmembran nachweisen, das Kennzeichen der basalen Seite eines Epithels.[33]
Der Ursprung des Kreislaufsystems wird also bei einem frühen bilateralen Tier vermutet, bei dem das Kreislaufsystem gegenwärtigen Vielborstern unter den Ringelwürmern ähnelte. Hier entwickelte sich eine Gruppe mesodermaler Zellen in ein Mesothel, das das Coelom auskleidete und Gefäße bildete. Mesotheliale Zellen differenzierten sich dann zu kontraktilen Gefäßzellen, Nephrozyten oder zu Blutvorläuferzellen. Derartige mesotheliale Zellen wären dann eine Art Vorläufer heutiger Hämangioblasten. Auch das Kreislaufsystem der Lanzettfischchen ähnelt dem des angenommenen ursprünglichen bilateralen Tiers.[33]
Wirbeltiere
Das Kreislaufsystem der Wirbeltiere unterscheidet sich davon. Die Gefäße sind von Endothel ausgekleidet und von Muskelzellen umgeben. Im Gegensatz zu den wirbellosen Coelomata trägt das Mesothel bei Wirbeltieren nicht zu den Gefäßen in den ausgewachsenen Tieren bei. Während der Embryogenese lässt sich die Verwandtschaft jedoch noch beobachten. Im frühen Embryo bildet das seitliche Mesoderm das Mesothel, das das Coelom umgibt. Das innere Blatt des Mesothels, die Splanchnopleura, bildet die Vorgängerzellen, die zu Endothel und Blutzellen werden (siehe AGM-Region)--> und auch die glatten Muskelzellen der Blutgefäßwände. Direkt neben der SplanchnopleuraZebrabärbling Danio rerio überlappen beide Regionen sogar.[33]
Die Vorläuferzellen der Blutgefäße und Blutzellen migrieren von der Splanchnopleura um die ersten Blutgefäße zu bilden. Endothelzellen beginnen mit der Bildung von Herz, Aorta, den großen Venen, sowie Verbindungen zwischen Aorta und Venen. Anschließend lagern sich mesodermale Zellen an, um die Muskelschicht aufzubauen. Blutstammzellen sondern sich im frühen Embryo vom Endothel der Aorta und anderer Gefäße ab. Weitere Blutstammzellen entstehen zumindest bei Säugetieren vermutlich im Dottersack und der Plazenta.[33]
Literatur
- Uwe Gille: Herz-Kreislauf- und Abwehrsystem, Angiologia. In: Franz-Viktor Salomon u. a. (Hrsg.): Anatomie für die Tiermedizin. 2., überarbeitete und erweiterte Auflage. Enke-Verlag, Stuttgart 2008, ISBN 978-3-8304-1075-1, S. 404–463.
- Rita Monahan-Earley, Ann M. Dvorak, William C. Aird: Evolutionary origins of the blood vascular system and endothelium. In: Journal of Thrombosis and Haemostasis.
Band 11, Supplement 1, Juni 2013, S. 46–66,
doi:
 10.1111/jth.12253,
PMC 5378490 (freier Volltext).
10.1111/jth.12253,
PMC 5378490 (freier Volltext).
Einzelnachweise
- ↑ Hochspringen nach: a b Neil A. Campbell, Jane B. Reece: Biologie. 8. Auflage. Pearson Deutschland, München 2009, ISBN 978-3-8273-7287-1, S. 1246.
- ↑ Hochspringen nach: a b
c d e
f g h
i j Rita Monahan-Earley, Ann M. Dvorak and William C. Aird:
Evolutionary origins of the blood vascular system and endothelium. In: J Thromb Haemost.
Band 11, Suppl 1, 2013,
S. 46–66,
doi: 10.1111/jth.12253,
PMC 5378490 (freier Volltext).
- ↑ Hochspringen nach: a b c d Christopher D. Moyes, Patricia M. Schulte: Tierphysiologie. Pearson Studium, München 2008, ISBN 978-3-8273-7270-3, S. 377–381 (Originaltitel: Principles of Animal Physiology. 2006.).
- ↑ Adolf Remane, Volker Storch, Ulrich Welsch: Kurzes Lehrbuch der Zoologie. 5. Auflage. Gustav Fischer Verlag, Stuttgart / New York, ISBN 3-437-20337-1, S. 184.
- ↑ Hochspringen nach: a b c d Heinz Penzlin: Lehrbuch der Tierphysiologie. 7. Auflage von 2005, unveränderter Nachdruck. Spektrum Akademischer Verlag, Heidelberg 2009, ISBN 978-3-8274-2114-2, S. 313.
- ↑ Christopher D. Moyes, Patricia M. Schulte: Tierphysiologie. Pearson Studium, München 2008, ISBN 978-3-8273-7270-3, S. 383–386 (Originaltitel: Principles of Animal Physiology. 2006.).
- ↑ Hochspringen nach: a b Adolf Remane, Volker Storch, Ulrich Welsch: Kurzes Lehrbuch der Zoologie. 5. Auflage. Gustav Fischer Verlag, Stuttgart 1985, ISBN 3-437-20337-1, S. 186–188.
- ↑ Hochspringen nach: a b c Heinz Penzlin: Lehrbuch der Tierphysiologie. 7. Auflage von 2005, unveränderter Nachdruck. Spektrum Akademischer Verlag, Heidelberg 2009, ISBN 978-3-8274-2114-2, S. 3550-351.
- ↑ Hochspringen nach: a b c d e f g h i j k l Gerhard Heldmaier, Gerhard Neuweiler, Wolfgang Rössler: Vergleichende Tierphysiologie. 2. Auflage. Springer Spektrum, Springer Verlag, Berlin/Heidelberg 2013, ISBN 978-3-642-25154-2, S. 258–262.
- ↑ Hochspringen nach: a b c Heinz Penzlin: Lehrbuch der Tierphysiologie. 7. Auflage von 2005, unveränderter Nachdruck. Spektrum Akademischer Verlag, Heidelberg 2009, ISBN 978-3-8274-2114-2, S. 349.
- ↑ Heinz Penzlin: Lehrbuch der Tierphysiologie. 7. Auflage von 2005, unveränderter Nachdruck. Spektrum Akademischer Verlag, Heidelberg 2009, ISBN 978-3-8274-2114-2, S. 333.
- ↑ Emile L. Boulpaep: The Micorcirculation. In: Walter F. Boron, Emile L. Boulpaep (Hrsg.): Medical Physiology. Third edition Auflage. Elsevier, Philadelphia, PA 2017, ISBN 978-1-4557-4377-3, S. 462.
- ↑ Rolf Kötter:
Wand der Leber-Sinusoide. (PDF) Anatomie.net, archiviert vom Internetarchiv
- ↑ A. Koo, I. Y. Lang: Microvascular filling pattern in rat liver sinusoids during vagal stimulation. In:
The Journal of Physiology. Band 295, Nr. 1, 1979, S. 191–199,
doi: 10.1113/jphysiol.1979.sp012961.
- ↑ Carl A. Goresky: A linear method for determining liver sinusoidal and extravascular volumes. In:
American Journal of Physiology-Legacy Content. Band 204, Nr. 4, April 1963, S. 626–640,
doi: 10.1152/ajplegacy.1963.204.4.626.
- ↑ Volker Storch, Ulrich Welsch: Systematische Zoologie. 6. Auflage. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-1112-2, S. 559–560.
- ↑ Rita Monahan-Earley, Ann M. Dvorak, William C. Aird: Evolutionary origins of the blood vascular system and
endothelium. In: Journal of Thrombosis and Haemostasis. Band 11, Supplement 1, Juni 2013, S. 46–66,
doi: 10.1111/jth.12253,
PMC 5378490 (freier Volltext).
- ↑ Christopher D. Moyes, Patricia M. Schulte: Tierphysiologie. Pearson Studium, München 2008, ISBN 978-3-8273-7270-3, S. 382 (Originaltitel: Principles of Animal Physiology. 2006.).
- ↑ Hochspringen nach: a b c Christopher D. Moyes, Patricia M. Schulte: Tierphysiologie. Pearson Studium, München 2008, ISBN 978-3-8273-7270-3, S. 383–384 (Originaltitel: Principles of Animal Physiology. 2006.).
- ↑ Heinz Penzlin: Lehrbuch der Tierphysiologie. 7. Auflage von 2005, unveränderter Nachdruck. Spektrum Akademischer Verlag, Heidelberg 2009, ISBN 978-3-8274-2114-2, S. 340–341.
- ↑ Volker Storch, Ulrich Welsch: Systematische Zoologie. 6. Auflage. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-1112-2, S. 238.
- ↑ Volker Storch, Ulrich Welsch: Systematische Zoologie. 6. Auflage. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-1112-2, S. 126.
- ↑ Hochspringen nach: a b c d e f g h Christopher D. Moyes, Patricia M. Schulte: Tierphysiologie. Pearson Studium, München 2008, ISBN 978-3-8273-7270-3, S. 384–386 (Originaltitel: Principles of Animal Physiology. 2006.).
- ↑ Georg Fischer: Chirurgie vor 100 Jahren. Historische Studie. Verlag von F. C. W. Vogel, Leipzig 1876; Neudruck mit dem Untertitel Historische Studie über das 18. Jahrhundert aus dem Jahre 1876 und mit einem Vorwort von Rolf Winau: Springer-Verlag, Berlin / Heidelberg / New York 1978, ISBN 3-540-08751-6, S. 289.
- ↑ Georg Fischer: Chirurgie vor 100 Jahren. Historische Studie. Verlag von F. C. W. Vogel, Leipzig 1876; Neudruck mit dem Untertitel Historische Studie über das 18. Jahrhundert aus dem Jahre 1876 und mit einem Vorwort von Rolf Winau: Springer-Verlag, Berlin / Heidelberg / New York 1978, ISBN 3-540-08751-6 , S. 289.
- ↑ Volker Storch, Ulrich Welsch: Systematische Zoologie. 6. Auflage. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-1112-2, S. 447–448.
- ↑ Hochspringen nach: a b Volker Storch, Ulrich Welsch: Systematische Zoologie. 6. Auflage. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-1112-2, S. 453–454.
- ↑ Margit Pavelka, Jürgen Roth: Functional Ultrastructure. An Atlas of Tissue Biology and Pathology. Springer, 2005, S. 232.
- ↑ Warren W. Burggren, Kjell Johansen: Circulation and respiration in lungfishes (Dipnoi). In:
Journal of Morphology. Band 190, Nr. S1, 1986, S. 217–236,
doi: 10.1002/jmor.1051900415.
- ↑ Herbert Reindell, Helmut Klepzig: Krankheiten des Herzens und der Gefäße. In: Ludwig Heilmeyer (Hrsg.): Lehrbuch der Inneren Medizin. Springer-Verlag, Berlin/Göttingen/Heidelberg 1955; 2. Auflage ebenda 1961, S. 450–598, hier: S. 454 f. (Der periphere Kreislauf).
- ↑ Paul Diepgen, Heinz Goerke: Aschoff/Diepgen/Goerke: Kurze Übersichtstabelle zur Geschichte der Medizin. 7., neubearbeitete Auflage. Springer, Berlin/Göttingen/Heidelberg 1960, S. 16.
- ↑ Volker Storch, Ulrich Welsch: Systematische Zoologie. 6. Auflage. Spektrum Akademischer Verlag, Heidelberg/Berlin 2004, ISBN 3-8274-1112-2, S. 80.
- ↑ Hochspringen nach: a b
c d e
f g h
i Volker Hartenstein und Lolitika Mandal: The blood/vascular system in a phylogenetic
perspective. In: BioEssays. Band 28,
Nr. 12, 2006,
S. 1203–1210,
doi: 10.1002/bies.20497.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 11.03. 2026