Art (Biologie)
Die Art, auch Spezies oder Species genannt, ist in der Biologie (einschließlich Virologie und Palichnologie) die Grundeinheit der Systematik. Jede biologische Art ist ein Resultat der Artbildung. Bislang gelang keine allgemeine Definition der Art, welche die theoretischen und praktischen Anforderungen aller biologischen Teildisziplinen gleichermaßen erfüllt. Vielmehr existieren in der Biologie verschiedene Artkonzepte, die zu unterschiedlichen Klassifikationen führen. Historisch wie auch aktuell spielen zwei Ansätze von Artkonzepten eine wichtige Rolle:
- auf der Grundlage einer angenommenen Fortpflanzungsgemeinschaft (biologischer Artbegriff)
- auf Grund der Annahme einer Gruppe von Individuen mit charakteristischen gemeinsamen Artmerkmalen, mithilfe derer die Zugehörigkeit zu einer Art klar abgrenzbar sein soll und die anhand äußerer (morphologischer Artbegriff) oder erblicher Merkmale bestimmt werden.
Mit dem Aufkommen der Kladistik ist seit den 1950er Jahren der auf dem biologischen Artbegriff beruhende, chronologisch definierte phylogenetische Artbegriff hinzugekommen, nach dem eine Art mit der Artspaltung, also der Bildung zweier Arten aus einer Ursprungsart, beginnt und mit ihrer erneuten Artspaltung oder aber ihrem Aussterben endet.
Das Problem der Artdefinition besteht aus zwei Teilproblemen:
- Gruppenbildung: Welche Populationen von Individuen gehören zusammen?
- Rangbildung: Welche der zahlreichen, ineinander geschachtelten Gruppen von Populationen mit abgestuften Ähnlichkeiten und Beziehungen können als „Art“ zusammengefasst werden?
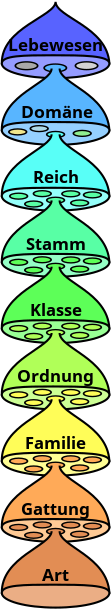
Die Hauptunterschiede der verschiedenen Artkonzepte liegen dabei auf der Ebene der Rangbildung. Eine Gruppe von Lebewesen unabhängig von ihrem Rang bezeichnen Taxonomen als Taxon (in der Botanik auch Sippe).
In biologischen Fachtexten wird bei Bezug auf eine unbestimmte Art oft abgekürzt „spec.“ oder „sp.“ (von lateinisch species), bei mehreren Arten auch „spp.“ (species pluralis).
Art als Taxon
Eine Art als Taxon ist eine gemäß den Regeln der Taxonomie und der biologischen Nomenklatur formal beschriebene und benannte Form von Lebewesen. Eine taxonomische Art stellt eine wissenschaftliche Hypothese dar und kann unabhängig von einem Artkonzept sein, sofern man zumindest akzeptiert, dass Arten reale und individuelle Erscheinungen der Natur sind. Die Art ist eine Rangstufe der klassischen, auf Carl von Linné zurückgehenden Taxonomie. Einige rein merkmalsbezogen arbeitende Systematiker sind der Ansicht, Arten wären mehr oder weniger willkürlich zusammengestellte, künstliche Gruppen, nur die Individuen seien letztlich real: Manche gehen dabei so weit, dass der Artbegriff wie alle anderen Rangstufen ihrer Ansicht nach besser abgeschafft werden sollten und durch neue Konzepte wie die Least-inclusive taxonomic unit ersetzt werden sollten. Die meisten Biologen sind aber der Ansicht, dass Arten natürliche Einheiten mit realer Existenz darstellen; es gäbe dann Artkriterien, an denen sich reale Arten identifizieren ließen. Dieser Vorstellung liegt letztlich eine Unterscheidung zwischen durch Genfluss oder horizontalem Gentransfer geprägten Einheiten unterhalb des Artniveaus und den Arten, bei denen dies nicht zutrifft (engl. lineages), zu Grunde. Für viele Biologen, darunter Anhänger eines phylogenetischen Artkonzepts (vgl. unten), sind sie sogar die einzigen in diesem Sinne natürlichen taxonomischen Einheiten.
Nomenklatur
Der wissenschaftliche Name einer Art (oft lateinischen oder griechischen Ursprungs) setzt sich nach der von Carl von Linné 1753 eingeführten binären Nomenklatur aus zwei Teilen zusammen, die beide kursiv geschrieben werden (diese Nomenklatur wurde 2021 auch in der Virologie für neue Virusarten eingeführt, die bestehenden anderweitigen Artnamen werden nach und nach umbenannt). Der erste Teil dieses Namens ist der groß geschriebene Gattungsname. Der zweite Teil wird immer klein geschrieben und in der Botanik sowie bei Prokaryoten als Epitheton („specific epithet“) bezeichnet, in der Zoologie als Artname oder Artzusatz („specific name“). Um Verwechslungen zwischen dem Artzusatz und dem gesamten Artnamen, also dem Binomen aus Gattungsname und Artzusatz, zu vermeiden, werden in der Zoologie entweder die eindeutigen englischen Begriffe verwendet oder hinzugefügt oder gelegentlich und informell auch Begriffe wie „epithetum specificum“ oder „epitheton specificum“ verwendet.
Beispiele
- Bei der Rotbuche (Fagus sylvatica) bezeichnet der Namensteil Fagus die Gattung, sylvatica ist das Artepitheton.
- Beim Löwen (Panthera leo) bezeichnet der Namensteil Panthera die Gattung, leo ist der Artname („specific name“).
Sowohl in der Botanik (Code Article 46) als auch in der Zoologie (Code Article 51) wird empfohlen, dem wissenschaftlichen Artnamen die Namen der Autoren beizufügen, welche die Art beschrieben haben, zumindest, wenn es um taxonomische oder nomenklatorische Fragen geht. Dies ist zum Beispiel wichtig, um Homonyme zu erkennen, das sind Fälle, in denen zwei Autoren versehentlich zwei verschiedene Arten mit demselben Namen benannt haben. Im Geltungsbereich des Internationalen Codes der Nomenklatur für Algen, Pilze und Pflanzen wird es empfohlen, die Autorennamen abzukürzen, wobei in der Regel das Namensverzeichnis von Brummit und Powell als Grundlage dient, „L.“ steht beispielsweise für Linné.
- Beispiel: Shiitake Lentinula edodes (Berk.) Pegler – Erklärung: Miles Josef Berkeley hat die Art zuerst beschrieben, David Norman Pegler hat sie in das heute gültige System eingeordnet.
Nach den Internationalen Regeln für die Zoologische Nomenklatur sollen zumindest einmal in jedem wissenschaftlichen Text dem Artnamen die Autor(en) und das Jahr der Publikation hinzugefügt werden (Code Recommendation 51a). Wenn im entsprechenden Fachgebiet zwei Autoren mit demselben Nachnamen tätig waren, soll der abgekürzte Vorname hinzugefügt werden, um Eindeutigkeit herzustellen. Wenn die Art heute in eine andere Gattung gestellt wird als in die, in der sie ursprünglich beschrieben wurde, müssen Autor(en) und Jahr in Klammern gesetzt werden (Code Article 51.3). Zwischen Autor und Jahr wird in der Regel ein Komma gesetzt.
- Beispiel: Löwe Panthera leo (Linnaeus, 1758) – Erklärung: Carl Nilsson Linnæus hat die Großkatze zuerst und als Felis leo beschrieben. Wer sie zuerst in die heute meist für den Löwen verwendete Gattung Panthera Oken, 1816 gestellt hat, ist in der Zoologie nicht relevant. Statt Linnæus wird Linnaeus geschrieben, da der Autor in lateinischen Buchstaben angegeben werden soll (Code Recommendation 51 b).
Geschichte
Die Philosophen der Antike kannten noch keine systematischen Konzepte und somit keinen Artbegriff im heutigen Sinne. Von Aristoteles sind als erstem Philosophen Schriften bekannt, in denen zwei getrennte – allgemein philosophisch zu verstehende – Begriffe είδος (eidos, ins Deutsche mit „Art“ übersetzt) und γένος (genos, deutsch „Gattung“) voneinander abgrenzt werden. In seinen Kategorien charakterisiert er anhand eines Beispiels aus der Welt der Lebewesen diese als zweite Wesenheiten (δεύτεραι ουσίαι), die in dem Einzelnen vorhanden sind. So ist ein einzelner Mensch in der Art Mensch vorhanden und ein einzelnes Pferd in der Art Pferd, beide gehören jedoch zur Gattung des Lebenden (ζῷον zoon).
In seiner Historia animalium (Περί τα ζώα ιστοριών) wendet Aristoteles die Begriffe είδος und γένος auch auf das Tierreich an, ohne dabei jedoch eine taxonomische Ordnung aufzustellen. Vielmehr spricht er von der Überlappung von Eigenschaften der Tierarten (ἐπάλλαξις epállaxis) und der Notwendigkeit, eine einzelne Art anhand mehrerer nebengeordneter Merkmale zu definieren. Dennoch beschäftigt er sich bei der Beschreibung der Arten mit einzelnen charakteristischen Merkmalen. Der Begriff είδος wird auch nicht im Sinne eines heutigen Artbegriffes konsequent als unterste Kategorie zwischen dem einzelnen Lebewesen und γένος verwendet, vielmehr kann die Bedeutung meist am besten mit „Form“, „Gestalt“ oder „Wesen“ wiedergegeben werden, während Tierarten in der Regel mit γένος bezeichnet werden.
Laut biblischer Schöpfungsgeschichte im 1. Buch Mose schuf Gott zwischen dem 3. und 6. Schöpfungstag die Pflanzen und Tiere, „ein jegliches (jedes) nach seiner Art“ (zehnmal Zitat „nach seiner Art“, Genesis 1,11–27 LUT, zu verstehen als „Wesensart“, hebräisch min מין bzw. למינה, Genesis 1,11–27 OT). In der Septuaginta wird מין mit γένος (κατὰ γένος „nach/gemäß der Art“, Genesis 1,11–27 LXX) übersetzt, in der Vulgata dagegen uneinheitlich, manchmal mit genus und manchmal mit species, wobei auch die Präpositionen wechseln (secundum speciem suam, secundum species suas, in species suas, juxta genus suum, secundum genus suum, in genere suo, Genesis 1,11–27 VUL). Es wird hier auch eine Aussage zur Fortpflanzung der Pflanzen und Tiere „nach ihrer Art“ getroffen, indem Gott in Genesis 1,11 LUT spricht: „Es lasse die Erde aufgehen Gras und Kraut, das Samen bringe, und fruchtbare Bäume auf Erden, die ein jeder nach seiner Art Früchte tragen, in denen ihr Same ist“, sowie in Genesis 1,22 LUT zu den Tieren des Wassers und der Luft: „Seid fruchtbar und mehret euch.“
Diese biblischen Aussagen wie auch Aristoteles waren bis in die Neuzeit prägend für die Vorstellungen der Gelehrten des Abendlandes. Pierre Duhem führte 1916 für die philosophische Auffassung vom Wesen oder der „Essenz“ eines Individuums den Begriff des Essentialismus ein.
Nach Auffassung von Ernst Mayr stimmte die auf dem „Schöpfungsglauben beruhende Interpretation des Artbegriffes der christlichen Fundamentalisten“ recht gut mit der letztendlich auf Platon zurückgehende Vorstellung einer „unveränderlichen Essenz“ (είδος als Wesen) überein und bildeten die Grundlage für einen „essentialistischen Artbegriff“, wie er vom Mittelalter bis ins 19. Jahrhundert hinein dominierte. Hiernach gehörten alle Objekte, welche dieselbe Essenz gemeinsam haben, derselben Art an. Laut Mayr war „[d]er Essentialismus mit seiner Betonung von Diskontinuität, Konstanz und typischen Werten (Typologie)“ der Hintergrund für typologische Artkonzepte, nach denen ein Individuum auf Grund seiner – in der Regel morphologischen – Merkmale (Typus) immer eindeutig einer bestimmten Art angehört und der Hintergrund dafür, dass Darwins „These von der Evolution durch natürliche Selektion daher als unannehmbar befunden“ wurde.
Erkennbar ist dies auch bei John Ray, der 1686 in seiner Historia plantarum generalis die Arten der Pflanzen als Fortpflanzungsgemeinschaften mit beständigen Artkennzeichen definiert, nachdem er „lange Zeit“ nach Anzeichen für ihre Unterscheidung geforscht habe: „Uns erschien aber keines [kein Anzeichen] zuverlässiger als die gesonderte Fortpflanzung aus dem Samen. Welche Unterschiede auch immer also im Individuum oder der Pflanzenart aus dem Samen derselben hervorgehen, sie sind zufällig und nicht für die Art kennzeichnend. […] Denn die sich nach ihrer Art unterscheiden, bewahren ihre Art beständig, und keine entspringt dem Samen der anderen oder umgekehrt.“
Systematisierung durch Carl von Linné
Carl von Linné stellte mit Species Plantarum (1753) und Systema Naturae (1758) als erster ein enkaptisches, auf hierarchisch aufbauenden Kategorien (Klasse, Ordnung, Gattung, Art und Varietät, jedoch noch nicht Familie) beruhendes System der Natur auf, wobei er für die Art die binäre Nomenklatur aus Gattungsnamen und Artepitheton einführte. Hierarchisch bedeutet dabei, dass die Einheiten auf unterschiedlichen Ebenen zu Gruppen zusammengefasst werden, wobei die in der Hierarchie höherstehenden Gruppen durch allgemeine, die tieferstehenden Gruppen durch immer speziellere Merkmale zusammengefasst werden (ein bestimmtes Individuum gehört also seiner Merkmalskombination gemäß in eine Art, eine Gattung, eine Familie usw.). Enkaptisch bedeutet, dass die in der Hierarchie tieferstehenden Gruppen in jeweils genau eine Gruppe der höheren Hierarchiestufe eingeschachtelt werden, also zum Beispiel jede Art in eine und genau eine, Gattung. In seiner Philosophia botanica formuliert er: „Es gibt so viele Arten, wie viele verschiedene Formen das unendliche Seiende am Anfang schuf; diese Formen, nach den hineingegebenen Gesetzen der Fortpflanzung, brachten viele [weitere Formen] hervor, doch immer ähnliche.“ Darüber hinaus bezeichnet er die Art und die Gattung als Werk der Natur, die Varietät als Werk des Menschen, Ordnung und Klasse dagegen als vom Menschen geschaffene Einheit. „Die Arten sind unveränderlich, denn ihre Fortpflanzung ist wahres Fortdauern.“
Während Georges-Louis Leclerc de Buffon 1749 noch verneint, dass es in der Natur irgendwelche Kategorien gäbe, revidiert er später diese Sicht für die Art und formuliert einen typologischen Artbegriff mit einer Konstanz der Arten: „Der Abdruck jeder Art ist ein Typ, dessen wesentliche Merkmale in unveränderlichen und beständigen Wesenszügen eingeprägt sind, doch alle Nebenmerkmale variieren: Kein Individuum gleicht vollkommen dem anderen.“
ean-Baptiste de Lamarck, der bereits von einer Transformation der Arten ausgeht, betrachtet dagegen die Art und alle anderen Kategorien als künstlich. 1809 äußert er sich in seiner Philosophie zoologique: „Die Natur hat nicht wirklich Klassen, Ordnungen, Familien, Gattungen, beständige Arten herausgebildet, sondern allein Individuen.“ Dies hindert ihn jedoch nicht daran, auf dem Gebiet der Taxonomie sehr produktiv zu sein, deren Kategorien er praktisch zu nutzen weiß.
Charles Darwin, der von der Art sogar im Titel seines Grundlagenwerkes On the Origin of Species (Über die Entstehung der Arten) von 1859 spricht, scheut sich vor einer Formulierung eines Artbegriffs. Laut Ernst Mayr kann man aus seinen Notizbüchern aus den 1830er Jahren schließen, dass er damals die Vorstellung von einer Art als Fortpflanzungsgemeinschaft hatte. In seiner Entstehung der Arten bezeichnet er jedoch die Begriffe der Art und der Varietät unmissverständlich als künstlich: „Aus diesen Bemerkungen geht hervor, dass ich den Kunstausdruck „Species“ als einen arbiträren und der Bequemlichkeit halber auf eine Reihe von einander sehr ähnlichen Individuen angewendeten betrachte und dass er von dem Kunstausdrucke „Varietät“, welcher auf minder abweichende und noch mehr schwankende Formen Anwendung findet, nicht wesentlich verschieden ist. Ebenso wird der Ausdruck „Varietät“ im Vergleich zu bloßen individuellen Verschiedenheiten nur arbiträr und der Bequemlichkeit wegen benutzt.“
Ähnlich äußert sich auch Alfred Russel Wallace 1856 in seiner Grundlagenarbeit über die Ritterfalter (Papilionidae) im Malaiischen Archipel, in der er verschiedene Verläufe der Evolution durch natürliche Zuchtwahl erklärt. Er bezeichnet Arten als „lediglich stark gekennzeichnete Rassen oder Lokalformen“ und geht dabei auch darauf ein, dass Individuen unterschiedlicher Arten generell als unfähig angesehen werden, fruchtbare gemeinsame Nachkommen zu zeugen, doch sei es nicht einmal in einem von tausend Fällen möglich, das Vorliegen einer Vermischung zu überprüfen.
Seit Darwin ist die Ebene der Art gegenüber unterscheidbaren untergeordneten (Lokalpopulationen) oder übergeordneten (Artengruppen bzw. höheren Taxa) nicht mehr besonders ausgezeichnet. Innerhalb der Taxonomie unterlag die Artabgrenzung Moden und persönlichen Vorlieben, es gibt Taxonomen, die möglichst jede unterscheidbare Form in den Artrang erheben wollen („splitter“), und andere, die weitgefasste Arten mit zahlreichen Lokalrassen und -populationen bevorzugen („lumper“).
Ende des 19. Jahrhunderts wurden biologische Artkonzepte einer Fortpflanzungsgemeinschaft diskutiert. Erwin Stresemann äußert in diesem Sinne bereits 1919 in einem Artikel über die europäischen Baumläufer klare Vorstellungen über Artbildung und genetische Isolation: „Es will nur die Tatsache im Namen zum Ausdruck bringen, dass sich die [im Laufe der geographischen Separation] zum Rang von Spezies erhobenen Formen physiologisch so weit voneinander entfernt haben, dass sie, wie die Natur beweist, wieder zusammenkommen können, ohne eine Vermischung einzugehen.“
Beherrschend im wissenschaftlichen Diskurs wurden die biologischen Artkonzepte der Fortpflanzungsgemeinschaft mit Theodosius Dobzhansky und Ernst Mayr seit der 2. Hälfte des 20. Jahrhunderts. Dobzhansky verknüpft den Artbegriff – ähnlich wie Stresemann – mit der Artbildung und definiert 1939 Arten als das „Stadium des Evolutionsvorgangs […], in dem Formengruppen, die sich bisher untereinander fortpflanzen oder jedenfalls dazu fähig waren, in zwei oder mehr gesonderte Gruppen aufgeteilt werden, die sich aus physiologischen Ursachen nicht untereinander fortpflanzen können“, während Mayr 1942 formuliert: „Arten sind Gruppen von natürlichen Populationen, die sich tatsächlich oder potentiell untereinander vermehren und fortpflanzungsmäßig von anderen derartigen Gruppen getrennt sind.“ In einem erweiterten biologischen Artbegriff bezieht Mayr 2002 die ökologische Nische mit in die Begriffsdefinition ein: „Eine Art ist eine Fortpflanzungsgemeinschaft von (fortpflanzungsmäßig von anderen isolierten) Populationen, die eine spezifische Nische in der Natur einnimmt.“ Mayr stellt die Bedeutung der Art in der Biologie als natürliche „Einheit der Evolution, der Systematik, der Ökologie und der Ethologie“ heraus und hebt sie hierin von allen anderen systematischen Kategorien ab.
Aus praktischen Erwägungen überdauern bis heute auch typologische Artkonzepte. Nach wie vor benennt die als Autorität bezeichnete Person, welche die Artbeschreibung einer neuen Art (species nova) als erste veröffentlicht, diese anhand der arttypischen Merkmale des Typusexemplars mit einem selbst gewählten Artnamen aus dem Gattungsnamen und dem Artepitheton.
Debatte um Essentialismus in der Geschichte der Biologie
In der Debatte um Essentialismus in der Geschichte der Biologie hebt Mary Winsor hervor, dass etwa die Verwendung von Typusarten als Prototypen für höhere Kategorien unvereinbar mit dem Essentialismus sei, und John S. Wilkins betont, dass die – von Winsor als „Methode der Exemplare“ bezeichnete – Typologie der Biologen und der Essentialismus keineswegs zwangsläufig verknüpft sind. Während Essenzen definierbar und allen Angehörigen einer Art eigen seien, ließen sich Typen instantiieren und seien variabel.
Laut Ernst Mayr beginnt die Geschichte des Artbegriffs in der Biologie mit Carl von Linné. Er hebt in seinen Arbeiten zur Wissenschaftsgeschichte hervor, dass der Essentialismus das abendländische Denken in großem Ausmaß beherrscht habe. Er setzt typologische mit essentialistischen Artbegriffen gleich, die mit Darwin's These von der Evolution durch natürliche Selektion „absolut unvereinbar“ sind.
„Darwin, einer der ersten Denker, der den Essentialismus (wenigstens zum Teil) ablehnte, wurde von den zeitgenössischen Philosophen (die alle Essentialisten waren) überhaupt nicht verstanden, und seine These von der Evolution durch natürliche Selektion daher als unannehmbar befunden. In essentialistischer Sicht ist eine echte Veränderung nur durch saltationistische, sprunghafte Entstehung neuer Wesenheiten möglich. Da die Evolution, wie Darwin sie erklärt, zwangsläufig allmählich, in fast unmerklichen Schritten erfolgt, ist sie mit dem Essentialismus absolut unvereinbar.“
Artkonzepte
Morphologisches Artkonzept
Typologisch definierte Arten sind Gruppen von Organismen, die in der Regel nach morphologischen Merkmalen (morphologisches Artkonzept) unterschieden werden. Es können aber auch andere Merkmale wie zum Beispiel Verhaltensweisen in analoger Weise verwendet werden. Eine nach morphologischen Kriterien definierte Art wird Morphospezies genannt.
Beispiele:
- Pferd und Esel lassen sich morphologisch klar voneinander abgrenzen und gehören damit zu verschiedenen Morphospezies.
- Löwe und
Tiger lassen sich morphologisch und im Verhalten klar voneinander
abgrenzen:
- Tiger sind gestreift und leben als Einzelgänger, die sich nur zur Paarungszeit treffen.
- Löwen haben nur als Jungtiere manchmal ein Fleckenmuster, sind nicht gestreift, die Männchen haben eine mehr oder weniger stark entwickelte Mähne. Löwen leben normalerweise in Rudeln aus Weibchen mit ihren Jungtieren, und einem oder mehreren adulten Männchen.
- Die Fellmerkmale und das Verhalten der Arten überlappen sich in ihrer Ausprägung nicht, und wenn (Liger und Tigons in Zoos), dann sind diese Zwischenformen viel seltener. Beides sind daher gut trennbare Morphospezies (bzw. Ethospezies).
In der Paläontologie kann in der Regel nur das morphologische Artkonzept angewandt werden. Da die Anzahl der Funde oft begrenzt ist, ist die Artabgrenzung in der Paläontologie besonders subjektiv. Beispiel: Die Funde von Fossilien zweier Individuen in der gleichen Fundschicht, also praktisch gleichzeitig lebend, unterscheiden sich stark voneinander:
- Sie können jetzt zwei verschiedenen Arten zugeordnet werden, wenn man der Meinung ist, dass sie weit genug von einem morphologischen Typus abweichen. Sie können aber auch derselben Art zugeordnet werden, wenn man der Meinung ist, dass in dieser Art auch eine größere Variationsbreite, welche die Funde mit einschließt, angenommen werden kann.
- Die Unterschiede können aber auch auf einen deutlichen Sexualdimorphismus (Unterschiede in der Erscheinung der Männchen und Weibchen) innerhalb einer Art zurückzuführen sein.
Diese Probleme werden mit zunehmender Zahl der Funde und damit Kenntnis der tatsächlichen Variationsbreite geringer, lassen sich aber nicht vollständig beseitigen.
Das morphologische Artkonzept findet häufig Verwendung in der Ökologie, Botanik und Zoologie. In anderen Bereichen, wie etwa in der Mikrobiologie oder in Teilbereichen der Zoologie, wie bei den Nematoden, versagen rein morphologische Arteinteilungsversuche weitgehend.
Problematik der morphologischen Abgrenzung
- Die Natur ist kein starres System, sondern in stetiger Veränderung begriffen. Unter dem Einfluss verschiedener Evolutionsfaktoren verändern sich Populationen graduell, gelegentlich auch sprunghaft von Generation zu Generation. Ein unveränderlicher Typus ist daher mit den Erkenntnissen der Evolutionsbiologie nicht vereinbar. In der belebten Natur gibt es keine Typen oder Essenzen (Ernst Mayr 1998).
- Eine Kategorisierung anhand morphologischer Merkmale ist nicht objektivierbar. Eine auf bloßer Unterscheidbarkeit basierende Einteilung hängt stets davon ab, wie genau man die verschiedenen Individuen oder Populationen untersucht und an welchen Kriterien die „Verschiedenheit“ festgemacht wird, was viel Raum für Willkür und Interpretation lässt. Je genauer die Untersuchungsmethoden, desto mehr Unterschiede zwischen verschiedenen Individuen und Populationen werden auffällig. In der Konsequenz würde jede noch so kleine intraspezifische Variation zu einem eigenen Taxon erklärt, wenn der jeweilige Taxonom den Unterschied für wesentlich erachtet. Durch die Existenz von Hybrid- und Übergangsformen wird das Problem zusätzlich verschärft, weil hier eine eindeutige, nicht willkürliche Abgrenzung nach morphologischen Gesichtspunkten kaum möglich ist.
- Der morphologische Artbegriff ist nicht konsequent durchzuhalten, weil er häufig im Widerspruch zur beobachtbaren biologischen Realität steht. In der Praxis ergibt sich diese Einschränkung u.a. aus der Existenz intraspezifischer Polymorphismen. Eine Reihe Spezies durchläuft während ihrer Individualentwicklung verschiedene Stadien (z.B. Larve → Fliege, Raupe → Schmetterling) in denen der jeweilige Phänotyp drastischen Veränderungen unterworfen ist. Häufig sind Sexualdimorphismen anzutreffen, Arten in denen männliche und weibliche Individuen unterschiedliche Phänotypen ausbilden. Beispielsweise ordnete Linné Männchen und Weibchen der Stockente ursprünglich zwei verschiedenen Arten zu; als man den Fehler erkannte, wurden beide zu einer Art zusammengefasst, obwohl sich an ihrer Unterschiedlichkeit nichts geändert hatte.
- Viele Spezies zeichnen sich durch eine hohe phänotypische Plastizität aus. Ein Phänotyp ist nicht vollständig durch den Genotyp determiniert, sondern das Ergebnis der Wechselwirkung von Genotyp und Umwelt. Derselbe Genotyp kann je nach Umwelt- und Lebensbedingungen unterschiedliche Standortformen hervorbringen, welche nach morphologischen Kriterien verschiedenen Taxa zugeordnet werden, obwohl sie genetisch völlig identisch sein können (z.B. im Falle von Ablegern). Beispielsweise variiert die Blattform des Löwenzahns sehr stark in Abhängigkeit von Niederschlagsmenge, Sonnenstrahlung und Jahreszeit zur Zeit der Blattbildung.
- Es gibt auch die umgekehrte Situation: Biologisch völlig verschiedene Arten können aufgrund ähnlicher Selektionsbedingungen in ihrem Phänotyp konvergieren, sodass sie rein äußerlich nicht mehr ohne Weiteres zu unterscheiden sind, so genannte Zwillingsarten. Das gleiche Problem stellt sich bei den kryptischen Arten.
- Schließlich erwies sich ein rein morphologisches Abgrenzungskriterium als nicht zuverlässig genug, weil die Variationen innerhalb einer Fortpflanzungsgemeinschaft größer sein können als diejenigen zwischen morphologisch ähnlichen, also Populationen desselben „Typus“, welche jedoch keine Fortpflanzungsgemeinschaft bilden.
Physiologisches Artkonzept bei Bakterien
Bakterien zeigen nur wenige morphologische Unterscheidungsmerkmale und weisen praktisch keine Rekombinationsschranken auf. In Ermangelung eindeutiger Abgrenzungsdefinitionen erstellte das International Committee on Systematics of Prokaryotes (ICSP) 2001 das weitest verbreitete Artenkonzept für Bakterien („Phylo-phenetic species concept“): „A monophyletic and genomically coherent cluster of individual organisms that show a high degree of overall similarity in many independent characteristics, and is diagnosable by a discriminative phenotypic property.“ (Ein monophyletisch und genomisch kohärentes Cluster einzelner Organismen, die in vielen unabhängigen Merkmalen einen hohen Grad an Gesamtähnlichkeit aufweisen und durch eine diskriminative phänotypische Eigenschaft diagnostizierbar sind.)
In der Praxis wird überwiegend der Stoffwechsel als Unterscheidungskriterium von Stämmen herangezogen. Weil ein allgemein akzeptiertes Artkriterium fehlt, stellen Bakterienstämme so die derzeit tatsächlich verwendete Basis zur Unterscheidung dar. Anhand biochemischer Merkmale wie etwa der Substanz der Zellwand unterscheidet man die höheren Bakterientaxa. Man testet an bakteriellen Reinkulturen zu ihrer „Artbestimmung“ deren Fähigkeit zu bestimmten biochemischen Leistungen, etwa der Fähigkeit zum Abbau bestimmter „Substrate“, z.B. seltener Zuckerarten. Diese Fähigkeit ist leicht erkennbar, wenn das Umsetzungsprodukt einen im Kulturmedium zugesetzten Farbindikator umfärben kann. Durch Verimpfung einer Bakterienreinkultur in eine Reihe von Kulturgläsern mit Nährlösungen, die jeweils nur ein bestimmtes Substrat enthalten („Selektivmedien“), bekommt man eine sog. „Bunte Reihe“, aus deren Farbumschlägen nach einer Tabelle die Bakterienart bestimmt werden kann. Dazu wurden halbautomatische Geräte („Mikroplatten-Reader“) entwickelt.
Seit entsprechende Techniken zur Verfügung stehen (PCR), werden Bakterienstämme auch anhand der DNA-Sequenzen identifiziert oder unterschieden. Ein weithin akzeptiertes Maß ist, dass Stämme, die weniger als 70 % ihres Genoms gemeinsam haben, als getrennte Arten aufzufassen sind. Ein weiteres Maß beruht auf der Ähnlichkeit der 16S-rRNA-Gene. Nach DNA-Analysen waren dabei zum Beispiel weniger als 1 % der in natürlichen Medien gefundenen Stämme auf den konventionellen Nährmedien vermehrbar. Auf diese Weise sollen in einem ml Boden bis zu 100.000 verschiedene Bakteriengenome festgestellt worden sein, die als verschiedene Arten interpretiert wurden. Dies ist nicht zu verwechseln mit der Gesamtkeimzahl, die in der gleichen Größenordnung liegt, aber dabei nur „wenige“ Arten umfasst, die sich bei einer bestimmten Kulturmethode durch die Bildung von Kolonien zeigen.
Viele Unterscheidungskriterien sind rein pragmatisch. Auf welcher Ebene der Unterscheidung man hier Stämme als Arten oder gar Gattungen auffasst, ist eine Sache der Konvention. Die physiologische oder genetische Artabgrenzung bei Bakterien entspricht methodisch dem typologischen Artkonzept. Ernst Mayr, leidenschaftlicher Anhänger des biologischen Artkonzepts, meint daher: „Bakterien haben keine Arten“.
Daniel Dykhuizen macht darauf aufmerksam, dass – entgegen mancher Anschauung – Transformationen, Transduktionen und Konjugationen (als Wege des DNA-Tauschs zwischen Stämmen) nicht wahllos, sondern zwischen bestimmten Formen bevorzugt, zwischen anderen quasi nie ablaufen. Demnach wäre es prinzipiell möglich, ein Artkonzept für Bakterien entsprechend dem biologischen Artkonzept bei den Eukaryonten zu entwickeln. Frederick M. Cohan versucht dagegen auf Basis von Ökotypen, ein Artkonzept zu entwickeln.
Biologisches oder populationsgenetisches Artenkonzept
Gegen Ende des 19., Anfang des 20. Jahrhunderts begann sich in der Biologie allmählich das Populationsdenken durchzusetzen, was Konsequenzen für den Artbegriff mit sich brachte. Weil typologische Klassifizierungsschemata die realen Verhältnisse in der Natur nicht oder nur unzureichend abzubilden vermochten, musste die biologische Systematik einen neuen Artbegriff entwickeln, der nicht auf abstrakter Unterschiedlichkeit oder subjektiver Einschätzung einzelner Wissenschaftler basiert, sondern auf objektiv feststellbaren Kriterien. Diese Definition wird als biologische Artdefinition bezeichnet, „Sie heißt „biologisch“ nicht deshalb, weil sie mit biologischen Taxa zu tun hat, sondern weil ihre Definition eine biologische ist. Sie verwendet Kriterien, die, was die unbelebte Welt betrifft, bedeutungslos sind.“ Eine biologisch definierte Art wird als Biospezies bezeichnet.
Der neue Begriff stützte sich auf zwei Beobachtungen: Zum einen setzen sich Arten aus Populationen zusammen und zum anderen existieren zwischen Populationen unterschiedlicher Arten biologische Fortpflanzungsbarrieren. „Die [biologische] Art besitzt zwei Eigenschaften, durch die sie sich grundlegend von allen anderen taxonomischen Kategorien, etwa dem Genus unterscheidet. Erstens einmal erlaubt sie eine nichtwillkürliche Definition – man könnte sogar so weit gehen, sie als „selbstoperational“ zu bezeichnen –, indem sie das Kriterium der Fortpflanzungsisolation gegenüber anderen Populationen hervorhebt. Zweitens ist die Art nicht wie alle anderen Kategorien auf der Basis von ihr innewohnenden Eigenschaften, nicht aufgrund des Besitzes bestimmter sichtbarer Attribute definiert, sondern durch ihre Relation zu anderen Arten.“ Das hat – zumindest nach der Mehrzahl der Interpretationen – zur Folge, dass Arten nicht Klassen sind, sondern Individuen.
Das Kriterium der Fortpflanzungsfähigkeit bildet den Kern des biologischen Artbegriffs oder der Biospezies. Eine Biospezies ist eine Gruppe sich tatsächlich oder potentiell miteinander fortpflanzender Individuen, die voll fertile Nachkommen hervorbringen:
- Eine Art ist eine Gruppe natürlicher Populationen, die sich untereinander kreuzen können und von anderen Gruppen reproduktiv isoliert sind.
Dabei sollen die Isolationsmechanismen zwischen den einzelnen Arten biologischer Natur sein, also nicht auf äußeren Gegebenheiten, räumlicher oder zeitlicher Trennung basieren, sondern Eigenschaften der Lebewesen selbst sein:
- Isolationsmechanismen sind biologische Eigenschaften einzelner Lebewesen, die eine Kreuzung von Populationen verschiedener sympatrischer Arten verhindern.
Die Kohäsion der Biospezies, ihr genetischer Zusammenhalt, wird durch physiologische, ethologische, morphologische und genetische Eigenschaften gewährleistet, die gegenüber artfremden Individuen isolierend wirken. Da die Isolationsmechanismen verhindern, dass nennenswerte zwischenartliche Bastardisierung stattfindet, bilden die Angehörigen einer Art eine Fortpflanzungsgemeinschaft; zwischen ihnen besteht Genfluss, sie teilen sich einen Genpool und bilden so eine Einheit, in der evolutionärer Wandel stattfindet.

Beispiele:
- Pferd und Esel sind zwar kreuzbar (Maultier, Maulesel), haben aber aufgrund einer genetischen Barriere keine fruchtbaren Nachkommen, bilden damit verschiedene Biospezies.
- Löwe und Tiger sind zwar unter künstlichen Bedingungen (Zoo) kreuzbar, (Großkatzenhybride: Liger Tigon) und haben im Zoo unter Umständen auch fruchtbare Nachkommen. In der Natur leben sie zwar teilweise in gemeinsamen Verbreitungsgebieten, natürliche Hybriden wurden bisher jedoch nicht nachgewiesen, was den Schluss nahelegt, dass sie sich nicht verpaaren. Sie gelten aufgrund ethologischer Barrieren als verschiedene Biospezies.
Problematik
- Geographisch deutlich getrennte Populationen sind, da sie sich in der Natur nicht kreuzen können, nach dem biologischen Artkonzept schwierig zu fassen. Nach der Theorie der allopatrischen Artbildung sind sie quasi „Arten im Entstehungsprozess“. Eine prinzipielle Schwierigkeit besteht eigentlich nicht, da die Frage experimentell entschieden werden kann (wenn keine biologischen Isolationsmechanismen evolviert sind, ist es noch dieselbe Art).
- Das biologische Artkonzept enthält in der ursprünglichen Fassung keinen Zeitbegriff. Untereinander kreuzen können sich evidenterweise nur gleichzeitig lebende Organismen. Ein Kriterium, ob früher lebende Organismen zur selben Art zu zählen sind oder nicht, wird dadurch nicht gegeben. Spätere Erweiterungen des Konzepts (zuerst wohl Simpson 1951) versuchten, dies durch Bezug auf evolutionär definierbare Einheiten zu überwinden.

- Arten, die sich nur ungeschlechtlich vermehren, werden durch die Definition des biologischen Artkonzepts nicht erfasst. Sie werden als Agamospezies bezeichnet. Hierzu gehören einige Protisten, einige kultivierte Form der Banane sowie einige Tiere mit parthenogenetischer Vermehrung). Agamospezies haben auch keinen Genpool und sind somit auch nach dem populationsgenetischen Artkonzept keine Arten.
- Viele Tier- und Pflanzenarten kreuzen sich auch in der Natur untereinander fruchtbar (Introgression), wie zum Beispiel verschiedene Steinkorallenarten oder Mehlbeer-Bäume sowie verschiedene Arten aus der Familie der Lebendgebärenden Zahnkarpfen jeweils innerhalb einer Gattung, wie beispielsweise Platy und Schwertträger in der Gattung Xiphophorus. Orchideen können sich zum Teil sogar über Gattungsgrenzen hinweg fruchtbar kreuzen. Diese Hybriden sind in der Natur in der Minderheit, die verschiedenen morphologisch beschriebenen Orchideenarten bleiben daher nach dem morphologischen Artkonzept unterscheidbar. Nach dem biologischen Artkonzept handelt es sich dann um getrennte Arten, wenn sich Isolationsmechanismen herausgebildet haben, die eine Hybridisierung normalerweise verhindern, auch wenn sie physiologisch möglich wäre, z.B. klimabedingte Unterschiede bei Tieren in der Fortpflanzungszeit oder bei Pflanzen in der Blütezeit. Diese Mechanismen können zusammenbrechen (z.B. durch menschliches Eingreifen oder drastische Änderungen der Umwelt durch Klimaveränderungen). Dadurch werden dem Konzept nach vorher getrennte Arten wieder zu einer Art (z.B. bei manchen Orchideenarten in Mitteleuropa beobachtet). Derselbe Vorgang kann aber auch natürlich ablaufen (introgressive Hybridisierung).
Das biologische Artkonzept findet häufig Verwendung in der Ökologie, Botanik und Zoologie, besonders in der Evolutionsbiologie. In gewisser Weise bildet es das Standardmodell, aus dem die anderen modernen Artkonzepte abgeleitet sind oder gegen welches sie sich in erster Linie abgrenzen. Die notwendigen Charaktere (Fehlen natürlicher Hybriden bzw. gemeinsamer Genpool) sind bisweilen umständlich zu überprüfen, in bestimmten Bereichen, wie etwa in der Paläontologie, versagen biologische bzw. populationsgenetische Artabgrenzungen weitgehend.
Phylogenetisches oder evolutionäres Artkonzept
Nach diesem Konzept wird eine Art als (monophyletische) Abstammungsgemeinschaft aus einer bis vielen Populationen definiert. Eine Art beginnt nach einer Artspaltung (Artbildung, Kladogenese) und endet
- wenn alle Individuen dieser Art, ohne Nachkommen zu hinterlassen, aussterben oder
- wenn aus dieser Art durch Artspaltung zwei neue Arten entstehen.
Phylogenetische Anagenese ist die Veränderung einer Art im Zeitraum zwischen zwei Artspaltungen, also während ihrer Existenz. Solange keine Aufspaltung erfolgt, gehören alle Individuen zur selben Art, auch wenn sie unter Umständen morphologisch unterscheidbar sind.
Das phylogenetische Artkonzept beruht auf der phylogenetischen Systematik oder „Kladistik“ und besitzt nur im Zusammenhang mit dieser Sinn. Im Rahmen des Konzepts sind Arten objektive, tatsächlich existierende biologische Einheiten. Alle höheren Einheiten der Systematik werden nach dem System „Kladen“ genannt und sind (als monophyletische Organismengemeinschaften) von Arten prinzipiell verschieden. Durch die gabelteilige (dichotome) Aufspaltung besitzen alle hierarchischen Einheiten oberhalb der Art (Gattung, Familie etc.) keine Bedeutung, sondern sind nur konventionelle Hilfsmittel, um Abstammungsgemeinschaften eines bestimmten Niveaus zu bezeichnen. Der wesentliche Unterschied liegt weniger in der Betrachtung der Art als in derjenigen dieser höheren Einheiten. Nach dem phylogenetischen Artkonzept können sich Kladen überlappen, wenn sie hybridogenen Ursprungs sind.
Problematik
- Jede Art und jede Artaufspaltung in diesem Modell muss zunächst, dem typologischen oder dem biologischen Artkonzept folgend, definiert werden. Dabei können die beim jeweiligen Artkonzept bereits besprochenen Schwierigkeiten auftreten. Das phylogenetische Artkonzept vereinfacht lediglich die Betrachtung zwischen zwei Artaufspaltungen, indem alle Populationen dieser Zeitspanne zu einer Art zusammengefasst werden. Ernst Mayr meint daher: „Es gibt nur zwei Art-Konzepte, alles andere sind Definitionen“
Zusätzlich kommen folgende Schwierigkeiten hinzu:
- Eine monophyletische Abstammungsgemeinschaft ist nicht unbedingt erkennbar. Der fehlende Nachweis morphologischer und genetischer Unterschiede kann eine bereits erfolgte Aufspaltung nicht ausschließen.
- Phylogenetische Aufgabelungen sind oft nicht symmetrisch und sind in einer der beiden abgespalteten Linien zuweilen ohne genetische und morphologische Folgen. Die Artgrenzen des phylogenetischen Artkonzepts können daher kaum nachvollziehbar zu bestimmten Zeitpunkten sich fertil kreuzende und morphologisch einheitliche Populationen trennen. Wenn eine kleine Gruppe einer Art von einem Kontinent auf eine Insel verfrachtet wird und dort z.B. aufgrund von starker Selektion schnelle Artbildung einsetzt – warum sollte dann aus den auf dem Kontinent zurückgebliebenen Lebewesen der Ursprungsart, die sich unter Umständen nicht oder nicht nachweisbar verändern, eine neue Art werden?
- Die Evolution vieler Taxa verläuft reticulat, das heißt vernetzt, und nicht linear sich aufgabelnd. Morphospezies und Biospezies können (zumindest in Einzelfällen) auf verschiedene Abstammungslinien zurückgehen und daher para- oder polyphyletisch sein.
Chronologisches Artkonzept
Ein weiterer Versuch, Arten in der Zeit klar abzugrenzen, ist das chronologische Artkonzept (Chronospezies). Auch hier wird die Art zunächst anhand eines anderen Artkonzepts definiert (meist das morphologische Artkonzept). Dann werden nach den Kriterien dieses Konzepts auch die Artgrenzen zwischen in einer Region aufeinanderfolgenden Populationen definiert. Dieses Konzept findet vorwiegend in der Paläontologie Anwendung und ist daher in der Regel eine Erweiterung des morphologischen Artkonzeptes um den Faktor Zeit:
- Eine Art wird durch eine Sequenz zeitlich aufeinander folgender Populationen charakterisiert, deren Individuen innerhalb einer bestimmten morphologischen Variationsbreite liegen.
Dieses Konzept ist dann gut anwendbar, wenn praktisch lückenlose Fundfolgen vorliegen.
Statistisches Artkonzept
In der Paläontologie, speziell in der Paläoanthropologie erweist sich die Zuordnung zu Arten und sogar die Zuordnung zu Gattungen allein anhand fossiler Knochen als schwierig. Anstelle einer kontravalenten Zuordnung wird daher von John Francis Thackeray eine wahrscheinlichkeitstheoretische Zuordnung vorgeschlagen. Anstelle der Frage, ob ein Fossil zur Spezies A und ein anderes zur Spezies B gehört, wird die Wahrscheinlichkeit, dass beide zur selben Spezies gehören, errechnet. Dazu wird eine möglichst große Reihe von Paaren unterschiedlicher morphometrischer Messpunkte von je zwei Individuen verglichen, bei denen die Artzugehörigkeit unsicher ist. Die Messwertpaare weichen stets voneinander ab. Sie streuen in Form einer Gaußschen Normalverteilung. Innerhalb dieser Verteilung wird definiert, in welchem Intervall um den Mittelwert (z.B. 2 Sigma) beide Individuen als derselben Art zugehörig betrachtet werden. Liegen die Messpunkte außerhalb des vorgegebenen Intervalls, werden die beiden Individuen als zwei verschiedene Arten betrachtet.
Artenzahl
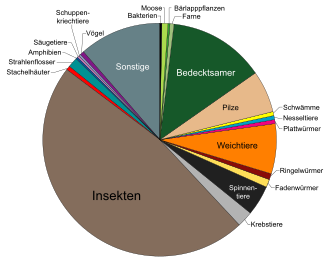
Anfang des 21. Jahrhunderts waren zwischen 1,5 und 1,75 Millionen Arten beschrieben, davon rund 500.000 Pflanzen. Es ist jedoch davon auszugehen, dass es sich bei diesen nur um einen Bruchteil aller existierenden Arten handelt. Schätzungen gehen davon aus, dass die Gesamtzahl aller Arten der Erde deutlich höher ist. Die weitestgehenden Annahmen reichten dabei Ende der 1990er-Jahre bis zu 117,7 Millionen Arten; am häufigsten jedoch wurden Schätzungen zwischen 13 und 20 Millionen Arten angeführt. Eine 2011 veröffentlichte Studie schätzte die Artenzahl auf 8,7 ± 1,3 Millionen, davon 2,2 ± 0,18 Millionen Meeresbewohner; diese Schätzung berücksichtigte allerdings nur Arten mit Zellkern (Eukaryoten), also nicht die Prokaryoten und auch nicht Viren, Viroide und Prionen.
Jay Lennon und Kenneth Locey von der Indiana University schätzten auf Basis der Ergebnisse von 3 Großprojekten, die Mikroben in Medizin, Meer und Boden behandeln, die Artenanzahl auf der Erde im März 2016 auf 1 Billion (1012). Insbesondere die kleinen Lebensformen der Bakterien, Archaeen und Pilze wurden bisher stark unterschätzt. Moderne Genom-Sequenzierung macht genaue Analysen möglich.
Über die Gesamtzahl aller Tier- und Pflanzenarten, die seit Beginn des Phanerozoikums vor 542 Mio. Jahren entstanden, liegen nur Schätzungen vor. Wissenschaftler gehen von etwa einer Milliarde Arten aus, manche rechnen sogar mit 1,6 Milliarden Arten. Weit unter einem Prozent dieser Artenvielfalt ist fossil erhalten geblieben, da die Bedingungen für eine Fossilwerdung generell ungünstig sind. Zudem zerstörten Erosion und Plattentektonik im Laufe der Jahrmillionen viele Fossilien. Forscher haben bis 1993 rund 130.000 fossile Arten wissenschaftlich beschrieben.
Es kann gezeigt werden, dass bei Verwendung des phylogenetischen Artkonzepts mehr Arten unterschieden werden als beim biologischen Artkonzept. Die Vermehrung der Artenzahl, z.B. innerhalb der Primaten, die ausschließlich auf das verwendete Artkonzept zurückgehen, ist als „taxonomische Inflation“ bezeichnet worden. Dies hat Folgen für angewandte Bereiche, wenn diese auf einem Vergleich von Artenlisten beruhen. Es ergeben sich unterschiedliche Verhältnisse beim Vergleich der Artenzahlen zwischen verschiedenen taxonomischen Gruppen, geographischen Gebieten, beim Anteil der endemischen Arten und bei der Definition der Schutzwürdigkeit von Populationen bzw. Gebieten im Naturschutz.
Literatur
- Neil A. Campbell: Biologie. Spektrum Akademischer Verlag, Heidelberg 1997, ISBN 3-8274-0032-5.
- Werner Kunz: Was ist eine Art? In der Praxis bewährt, aber unscharf definiert. In: Biologie in unserer Zeit. Wiley-VCH, Weinheim 32, 1, 2002, S. 10–19. ISSN 0045-205X
- Ernst Mayr: Das ist Leben – die Wissenschaft vom Leben. Spektrum Akademischer Verlag, Heidelberg 1997, ISBN 3-8274-1015-0.
- Ernst Mayr: Animal Species and Evolution. 6. Auflage. Belknap of Harvard University Press, Cambridge 1963, 1977; Artbegriff und Evolution. Parey, Hamburg/ Berlin 1967 (deutsch).
- Ernst Mayr: Grundlagen der zoologischen Systematik. Blackwell Wissenschaftsverlag, Berlin 1975, ISBN 3-490-03918-1.
- Ernst Mayr: Evolution und die Vielfalt des Lebens. Springer-Verlag, 1979, ISBN 3-540-09068-1.
- Peter Ax: Das Phylogenetische System. Urban & Fischer Bei Elsevier, 1997, ISBN 3-437-30450-X.
- Peter Ax: Systematik in der Biologie. Verlag Gustav Fischer, Stuttgart 1988, ISBN 3-437-20419-X.
- Peter Heuer: Art, Gattung, System: Eine logisch-systematische Analyse biologischer Grundbegriffe. Verlag Karl Alber, Freiburg i. Br. 2008, ISBN 978-3-495-48333-6.
- Robert A. Wilson (hrsg.): Species – New Interdisciplinary Essays. The MIT Press, Cambridge, Massachusetts, London 1999, ISBN 0-262-23201-4.
- Elliott Sober: Philosophy of Biology. 2. Auflage. Westview Press, 2000, ISBN 0-8133-9126-1.
- Rainer Willmann: Die Art in Raum und Zeit. Das Artkonzept in der Biologie und Paläontologie. Parey, Hamburg 1985, ISBN 3-489-62134-4.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 08.04. 2026