T-Gedächtniszelle
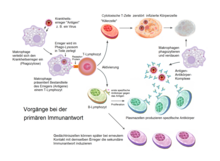
T-Gedächtniszellen oder Gedächtnis-T-Zellen sind eine Subpopulation (Untergruppe) der T-Lymphozyten (T-Zellen). Sie haben die Funktion eines immunologischen Gedächtnisses und verbessern den Schutz eines Individuums bei einer erneuten Infektion (Reinfektion) mit demselben Pathogen (Krankheitserreger). Als ehemalige T-Helferzellen (TH1 oder TH2) haben die Gedächtniszellen die spezifische, einmal erworbene Immunantwort gespeichert. Wenn der Organismus wieder mit demselben Antigen konfrontiert wird, lösen die T-Gedächtniszellen eine schnelle und effektive Immunreaktion aus. Teilweise wandeln sie sich dann erneut in T-Helferzellen um.[1][2][3]
Geschichte
Die Existenz von Gedächtniszellen konnte durch epidemiologische Beobachtungen bei einer Masernepidemie im 19. Jahrhundert auf den Färöer-Inseln angenommen werden. Damals fand man nach einmaliger Virus-Exposition und offensichtlich fehlender erneuter Exposition in der dazwischenliegenden Zeit noch nach 30 und mehr Jahren einen Schutz vor Neuerkrankung bei den ursprünglich an Masern erkrankten Bewohnern.[4]
Eigenschaften
Nach Stimulation des adaptiven Immunsystems durch ein Antigen erfolgt eine 10- bis 100fache Vermehrung der spezifischen T-Zellen. Von diesen können einige bei erneutem Kontakt mit dem gleichen Antigen eine schnellere und effizientere Sekundärantwort vermitteln. Diese Erinnerungsfunktion kann durch CD4-positive und durch CD8-positive-T-Gedächtniszellen übernommen werden.
Die aus naiven T-Zellen durch Differenzierung entstandenen Effektorzellen – ausdifferenzierte Lymphozyten mit spezifischen Aufgaben im Rahmen der Immunantwort – die meist nur eine kurze Lebenszeit haben und nach einer Immunantwort durch einen aktivierungs-induzierten Zelltod (engl. activation-induced cell death, AICD – programmierter Zelltod nach Aktivierung von T-Zellen und B-Zellen) absterben. An diesem Programm sind verschiedene Mechanismen beteiligt, unter anderem die Bereitstellung von Fas-vermittelten Signalen (CD95), intrazelluläre pro-apoptotischen Moleküle und der Verlust an Wachstumsfaktoren.
Von diesem programmierten Zelltod der während der spezifischen Immunantwort gebildeten T-Gedächtniszellen werden weniger als 5 % aller Effektorzellen ausgenommen. Die zellulären und molekularen Einflüsse, die darüber bestimmen, dass sich infolge einer Aktivierung naive T-Zellen zum einen in kurzlebige Effektorzellen und zum anderen in langlebige Gedächtniszellen differenzieren, sind nur in Ansätzen bekannt. Folgende Modelle zur Differenzierung von T-Gedächtniszellen werden diskutiert:
- Ein Modell setzt die Existenz von zwei verschiedenen naiven T-Zell-Untergruppen voraus, von denen eine das Potential besitzt, zu T-Effektorzellen auszureifen, während sich die andere nur zu T-Gedächtniszellen entwickeln kann. Überzeugende experimentelle Daten, die dieses Modell stützen könnten, fehlen zurzeit.
- In einem anderen Modell bestimmen unterschiedliche Signale bei der Aktivierung von naiven T-Zellen darüber, ob eine Effektorzelle oder eine T-Gedächtniszelle gebildet wird. Kostimulator-vermittelte Signale übermitteln, ob eine kurzlebige Effektorzelle als Folge starker Aktivierung oder eine langlebige Gedächtniszelle nach schwachen Signalen entstehen soll. Dieses Modell wird durch tierexperimentelle Beobachtungen unterstützt.
- Im dritten Modell überlebt eine Untergruppe von aktivierten Effektorzellen als T-Gedächtniszellen, weil sie entweder Signale erhalten, welche diese Differenzierung festhalten, oder weil sie sich den Signalen entziehen, die sonst den programmierten Zelltod einleiten. Offen bleibt, ob diese zusätzlichen Reize zufällig (stochastisch) auf die T-Effektorzellen wirken oder durch eine selektive Bereitstellung nur auf bestimmte Zellen wirken. Tierexperimentelle Ergebnisse sind mit diesem Modell vereinbar.
Typen
Wie bei naiven T-Zellen unterscheidet man bei den Gedächtnis-T-Zellen CD4- beziehungsweise CD8-positive Zellen. Darüber hinaus unterscheidet man innerhalb dieser zwei Typen mindestens zwei funktionell unterschiedliche Gruppen. Dies sind zum einen Effektor-Gedächtnis-T-Zellen (engl. effector memory T cells, TEM) und zum anderen zentrale Gedächtnis-T-Zellen (engl. central memory T cells, TCM). Beide Gruppen können sowohl bei CD4- als auch bei CD8-positiven Gedächtnis-T-Zellen vorkommen. Die Effektor-Gedächtnis-T-Zellen werden bei erneutem Antigenkontakt reaktiviert und stellen durch mehrere Teilungen viele funktionsfähige Effektorzellen bereit. Die zentralen Gedächtnis-T-Zellen ruhen dagegen in den Sekundären Lymphatischen Organen und haben zunächst keine Effektorfunktion am Infektionsherd. Allerdings können sie leicht aktiviert werden um sich zu Effektor-Zellen zu entwickeln, sobald das Antigen erneut auftritt. Die Arbeitsteilung zwischen Effektor-Gedächtnis-T-Zellen und zentralen Gedächtnis-T-Zellen besitzt ähnliche Züge wie die zwischen Plasma-Zellen und Gedächtnis-B-Zellen.[5]
- Effektor-Gedächtnis-T-Zellen des Menschen werden phänotypisch dadurch charakterisiert, dass sie kein CCR7 und in der Regel kein CD62L exprimieren, zwei Marker, die typischerweise auf naiven T-Zellen exprimiert werden. Effektor-Gedächtnis-T-Zellen können besonders schnell durch ihr Antigen aktiviert werden und sezernieren dann innerhalb weniger Stunden IFN-γ, IL-4, und IL-5.[5]
- Zentrale Gedächtnis-T-Zellen besitzen neben dem Gedächtnis-T-Zell-Marker CD45R0 (naive T-Zellen exprimieren CD45RA) auch CCR7 und CD62L. Diese Zellen sind leichter aktivierbar als naive T-Zellen und benötigen weniger Kostimulation, um aktiviert zu werden. Sie regulieren die Expression von CD40L auf ihrer Zelloberfläche hoch, wodurch sie leichter von dendritischen Zellen oder B-Zellen aktiviert werden können. Nach Aktivierung des T-Zell-Rezeptors produzieren diese Zellen vor allem Interleukin-2, nach einer ersten Vermehrungsphase differenzieren sie in Effektorzellen, die ebenfalls IFN-γ (bei TH1) oder IL-4 (bei TH2) produzieren.[5]
Literatur
- G. A. Holländer: Immunologie, Grundlagen für Klinik und Praxis. 1. Auflage. Elsevier, München 2006, ISBN 3-437-21301-6.
- M. J. Owen, J. R. Lamb: Immunerkennung. Thieme, Stuttgart 1991, ISBN 3-13-754101-8.
- Nicole Menche, Bernd Guzek, Hubert Hasel, Herbert Renz-Polster: Biologie Anatomie Physiologie. 6. Auflage. München 2007, ISBN 978-3-437-26801-4.
Einzelnachweise
- ↑ R. D. Michalek, J. C. Rathmell: The metabolic life and times of a T-cell. In:
Immunol Rev. (2010), Band 236, S. 190–202.
 PMID 20636818;
PMC 2983473 (freier Volltext).
PMID 20636818;
PMC 2983473 (freier Volltext).
- ↑ K. K. McKinstry, T. M. Strutt, S. L. Swain: Hallmarks of CD4 T cell immunity against influenza. In:
J Intern Med. (2011), Band 269(5), S. 507–518.
doi:10.1111/j.1365-2796.2011.02367.x.
PMID 21362069;
PMC 3395075 (freier Volltext).
- ↑ R. Arens, S. P. Schoenberger: Plasticity in programming of effector and memory CD8 T-cell formation.
In: Immunol Rev. (2010), Band 235(1), S. 190–205.
PMID 20536564;
PMC 2937176 (freier Volltext).
- ↑ Haven Emerson: Panum on Measles: Observations Made During the Epidemic of Measles on the Faroe Islands in
the Year 1846. In: Am J Public Health Nations Health. 1940, Band 30, Nummer 10, S. 1245–1246. (Übersetzung eines Artikel von Peter Ludwig Panum von 1938 aus dem
Dänischen) PMC 1530953 (freier Volltext)
- ↑ Hochspringen nach: a b c Federica Sallusto, Jens Geginat, Antonio Lanzavecchia: Central Memory and Effector Memory T cell Substets: Function, Generation, and Maintenance. In: Annu Rev Immunol. (2004). Band 22, S. 745–763.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 02.04. 2026