Diplohaplont
Diplohaplonten oder Haplo-Diplonten sind Lebewesen, bei deren Fortpflanzung abwechselnd haploide und diploide Generationen auftreten. Diese Art des Generationswechsels bezeichnet man als heterophasisch (griech. ἕτερος heteros ‚der andere‘, ‚ungleich‘), weil der Ploidiegrad (die Kernphase) der Generationen verschieden ist (vgl. Kernphasenwechsel). Zu den Diplohaplonten gehören alle „höheren“ Pflanzen (Embryophyta oder Landpflanzen) und die meisten Algen, von den Tieren hingegen nur die einzelligen Foraminiferen.
Kennzeichnend für Diplohaplonten ist, dass sowohl in der haploiden wie in der diploiden Phase Mitosen erfolgen, d.h. Kernteilungen ohne Änderung der Kernphase. Die haploide Generation pflanzt sich geschlechtlich fort, indem sie weibliche und männliche Geschlechtszellen (Gameten) bildet. Sie wird deshalb als Gametophyt bezeichnet. Durch die Vereinigung zweier Gameten verschiedenen Geschlechts und Verschmelzung der Zellkerne (Befruchtung) entsteht eine diploide Zygote, die erste Zelle der diploiden Generation. Diese pflanzt sich ungeschlechtlich fort, indem sie nach einer Reduktionsteilung (Meiose) haploide Sporen bildet. Sie wird deshalb als Sporophyt bezeichnet. Aus den Sporen wächst die nächste haploide Generation.
Sowohl der Gametophyt als auch der Sporophyt sind mehrzellig, aber die beiden Generationen haben meist völlig verschieden aussehende Erscheinungsformen. Von den Diplohaplonten zu unterscheiden sind Haplonten, bei denen nur die Zygote diploid ist – zu diesen gehören manche Algen und tierische Einzeller –, und Diplonten, bei denen nur die Gameten haploid sind – zu diesen gehören die meisten Tiere und der Mensch.
Ein nur etymologisch ähnlicher Begriff ist Haplodiploidie, eine Form der Geschlechtsdetermination.
Forschungsgeschichte
Dass alle höheren Pflanzen oder Landpflanzen einen Lebenszyklus mit zwei abwechselnden vielzelligen Generationen haben, beschrieb erstmals Wilhelm Hofmeister 1851. Ladislav Josef Čelakovský erkannte, dass der Sporophyt phylogenetisch jünger ist als der Gametophyt. Er stellte 1874 die beiden Generationen als gegensätzlich dar, weil sie morphologisch verschieden sind, sich unterschiedlich entwickeln und unterschiedliche Funktionen im Lebenszyklus haben. Eine konträre (nach heutiger Kenntnis falsche) Ansicht vertrat 1878 Nathanael Pringsheim, indem er die beiden Generationen als homolog betrachtete und die Verhältnisse bei den Moosen, bei denen der Gametophyt dem Sporophyten untergeordnet ist, als phylogenetisch abgeleitet interpretierte. Zu dieser Zeit hatte man die sexuelle Fortpflanzung noch nicht verstanden. Der Vorgang der Befruchtung war zwar bekannt, aber den komplementären Vorgang der Meiose klärte erst 1883 Édouard van Beneden auf. Erst dadurch wurde klar, dass sich der Generationswechsel bei Pflanzen nicht mit dem bei Tieren gleichsetzen lässt, weil dort kein Kernphasenwechsel erfolgt. Wie die evolutionär neue diploide Generation entstanden war, erklärte Frederick Orpen Bower im Jahr 1908: durch eine Verzögerung der Meiose, indem in der diploiden Phase Mitosen auftraten.[1]
Landpflanzen (Embryophyta)
Übersicht
Lebenszyklen verschiedener Landpflanzen:
-
 Lebermoose
Lebermoose -
 Laubmoose
Laubmoose -

 Hornmoose
Hornmoose -
Farne -

Bedecktsamer
Moose
-
 Gametophyten und Sporophyten des Lebermooses Pellia epiphylla
Gametophyten und Sporophyten des Lebermooses Pellia epiphylla -
 Sporenkapsel des Laubmooses Physcomitrella patens
Sporenkapsel des Laubmooses Physcomitrella patens -
 Protonema von Physcomitrella
Protonema von Physcomitrella
Bei den Leber- und Laubmoosen ist die haploide Generation, der Gametophyt, die äußerlich dominante „eigentliche“ grüne Moospflanze. Er liegt in zwei Geschlechtern vor, männlich und weiblich, und bildet Geschlechtsorgane (Gametangien), die im weiblichen Geschlecht als Archegonien bezeichnet werden und die Eizelle enthalten. Die männlichen Gametangien werden Antheridien genannt; in ihnen entstehen zweigeißelige Spermatozoide. Für die Befruchtung ist Wasser notwendig, denn die männlichen Gameten müssen zu den weiblichen schwimmen.
Die diploide Zygote entwickelt sich zu einem Embryo und weiter zum Sporophyten. Dieser bleibt bei Moosen mit dem Gametophyten verbunden und erhält von ihm Wasser und Nährstoffe. Er wächst durch das Archegonium hindurch und bildet meist einen Stiel (die Seta), an dessen Spitze eine Sporenkapsel (Sporangium) sitzt. Alternativ kann auch vom Gametophyten ein Stiel gebildet werden, der dann Pseudopodium genannt wird und an seiner Spitze den Sporophyten trägt. In der Sporenkapsel bilden sich durch Meiose haploide Sporen. Der Stiel ermöglicht eine bessere Ausbreitung der Sporen durch den Wind. Aus Sporen, die zu Boden fallen, wächst zunächst ein fädiger Vorkeim, das Protonema. An diesem bilden sich aus Knospen die haploiden Moospflanzen, und der Kreislauf ist geschlossen.
Bei Hornmoosen ist der haploide Gametophyt ein lappiger, flacher Thallus. Es gibt kein Protonema. Nach der Befruchtung wächst der Sporophyt horn- oder schotenförmig aus dem Gametophyten heraus. Er ist grün und treibt Photosynthese, bleibt aber wie bei den anderen Moosen von der Mutterpflanze abhängig. Aus Sporen, die der Sporophyt bildet, wächst die nächste haploide Generation heran.
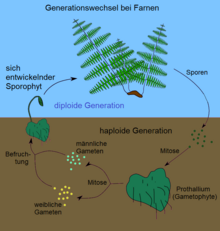
Farne

Bei den Farnen ist umgekehrt der diploide Sporophyt die dominierende Generation. An der Unterseite von Farnwedeln werden nach der Meiose haploide Sporen gebildet, die zu Boden fallen bzw. durch den Wind verbreitet werden und unter geeigneten Bedingungen die haploide Generation hervorbringen. Diese wird bei Farnen Prothallium genannt und ähnelt dem Thallus der Hornmoose. Sie bleibt deutlich kleiner als die diploide Farnpflanze, ausgewachsen meist nur ein bis zwei Zentimeter, und pflanzt sich geschlechtlich fort (Gametophyt). Auch hier ist für die Befruchtung Wasser notwendig, durch das die Spermatozoiden zu den Eizellen gelangen können.
Samenpflanzen

Auch bei den Samenpflanzen dominieren die Sporophyten die Gametophyten. Die „eigentliche“ Pflanze ist die Sporophyten-Generation, während die Gametophyten nur wenige Zellen groß sind.
Auf der diploiden Pflanze wachsen Fortpflanzungsorgane in Form von Blüten mit Fruchtblättern und Staubblättern. In diesen bilden sich durch Meiose haploide Zellen. Aus diesen wächst quasi auf der Pflanze die haploide Generation, die aber winzig klein bleibt. Der weibliche Gametophyt ist der meist nur aus wenigen Zellen bestehende Embryosack in der Samenanlage. Die männlichen Gametophyten sind die auf den Staubblättern gebildeten Mikrosporen, die zum haploiden mehrzelligen Inneren von Pollenkörnern auswachsen. Die Befruchtung ist anders als bei Farnen und Moosen vom Wasser unabhängig. Bei der Bestäubung wird zunächst das Pollenkorn auf das Fruchtblatt übertragen. Dort bildet es einen Pollenschlauch aus, der in das Fruchtblatt zur pflanzlichen Eizelle hinein wächst. Dort findet die eigentliche Befruchtung statt.
Bei der Befruchtung bildet sich die diploide Zygote. Diese wächst zunächst auf der Elternpflanze zu einem diploiden Samen heran. Dann legt der Same eine Wachstumspause ein, in der er Perioden großer Trockenheit und Kälte ohne Schaden überstehen kann. Anschließend kommt der Same unter günstigen Umweltbedingungen (Wärme und Feuchtigkeit) zur Keimung. Aus dem Keim entwickelt sich durch Mitosen die neue Pflanze.
Die Samenpflanzen können entweder einhäusig oder zweihäusig sein. Bei den einhäusigen gibt es auf derselben Pflanze sowohl weibliche als auch männliche Blüten. Die Blüten selbst sind eingeschlechtig, haben also entweder nur (männliche) Staubblätter oder nur (weibliche) Fruchtblätter. Bei zweihäusigen Samenpflanzen kommen weibliche und männliche Blüten nur auf getrennten Individuen vor. Daneben gibt es „echt zwittrige“ Pflanzen, die nur eine Art von Blüten haben, in denen sich gleichzeitig männliche und weibliche Geschlechtsorgane befinden.
Evolution
Die nächsten Verwandten der Embryophyta, die Armleuchteralgen, sind Haplonten, also mit Ausnahme der Zygote haploid. Dagegen bilden alle Embryophyta vielzellige diploide Sporophyten. Diese sind bei den ursprünglichsten Embryophyten, den Lebermoosen und Laubmoosen, nicht zur Photosynthese fähig und daher wie die Zygote der Armleuchteralgen von der Ernährung durch die Mutterpflanze (den Gametophyten) abhängig. Bei den Hornmoosen, die näher mit den Gefäßpflanzen verwandt sind als die anderen Moose, sind die Sporophyten photosynthetisch aktiv (was bei einigen Laubmoosen nur in geringem Umfang auftritt), bleiben aber auf die Versorgung durch die Mutterpflanze angewiesen. Bei den Gefäßpflanzen (Bärlapppflanzen, Farne und Samenpflanzen) schließlich ist der Sporophyt ein autarker, frei lebender Organismus, und bei den Samenpflanzen ist umgekehrt der stark reduzierte Gametophyt zur vollständigen Ernährung durch die Mutterpflanze übergegangen. Ein Übergangsstadium zwischen den Moosen und den heutigen Gefäßpflanzen mit wohlentwickelten Gametophyten und Sporophyten zeigen manche frühe Landpflanzen, die nur noch fossil erhalten sind.[1]
Der frei lebende Sporophyt der Gefäßpflanzen unterscheidet sich in mehrfacher Hinsicht sehr von dem auf dem Gametophyten sitzenden Sporophyten der Moose. Grundlegend ist die Ausbildung einer Wurzel als neuartiges Organ, ohne das die Selbständigkeit nicht möglich wäre. Dazu gehört die bei den Moosen fehlende oder schwach ausgeprägte Fähigkeit, auf die Schwerkraft zu reagieren (Gravitropismus), wobei die Wurzel positiv gravitrop der Schwerkraft folgt, während der Spross senkrecht nach oben wächst (negativ gravitrop). Neuartig sind außerdem die seitlich an der Sprossachse sitzenden Blätter. Die Vergrößerung und Verselbstständigung des Sporophyten ging einher mit einer Reduktion und Internalisierung (Verinnerlichung) des Gametophyten. Letztere begann schon bei den Hornmoosen, indem die Archegonien im Thallus versanken. Hinzu kam die Heterosporie, eine zunehmende Spezialisierung der männlichen und weiblichen Sporen.[1]
Das Sporangium der Moose ist eine evolutionäre Neuentwicklung, also nicht homolog mit den ebenso bezeichneten Organen bei Algen. Es besteht aus sporogenem (sporenbildendem) Gewebe und einer Hülle. Es ist das einzige Organ, das bei den Sporophyten aller Embryophyta vorhanden ist. Bei den Moosen besteht der Sporophyt nur aus einem Sporangium und einem Stiel. Bei den Samenpflanzen sind die Sporangien als Pollensäcke an den Staubblättern und als Nucellus in der Samenanlage differenziert.[1]
Algen
Die meisten Grünalgen sind Haplonten, es gibt jedoch auch Diplohaplonten. Bei letzteren kann der Generationswechsel isomorph sein, d.h. die haploide und die diploide Generation sind äußerlich gleich, etwa beim Meersalat, oder heteromorph, wobei die diploide Generation in den Vordergrund tritt.[2]
Viele, aber nicht alle Braunalgen sind Diplohaplonten und vollziehen einen heterophasischen Generationswechsel. Ursprünglich war dieser Generationswechsel isomorph, wie etwa bei den meisten Arten der Gattung Ectocarpus. Daraus entwickelten sich Braunalgen mit heteromorphem Generationswechsel, bei denen die beiden Generationen unterschiedliches Aussehen haben. Bei den Fucales ist die haploide Generation so weit rückgebildet, dass sie fast reine Diplonten sind, sie gelten nicht mehr als Diplohaplonten. Viele Algen bilden Sporen, die begeißelt und deshalb im Wasser frei beweglich sind. Sie werden Zoosporen oder Schwärmsporen genannt.
Die Rotalgen sind Diplohaplonten mit einem einzigartigen dreigliedrigen Generationswechsel: Eine haploide Generation besteht aus männlichen und weiblichen Gametophyten. Die männlichen Gameten sind unbegeißelt und werden im Wasser passiv zu den weiblichen Gameten getrieben, die sich in einem Gametangium des weiblichen Gametophyten befinden. Dort kommt es zur Befruchtung. Die diploide Zygote wächst direkt auf dem Gametophyten zur zweiten Generation heran, die meist nur aus unscheinbaren, mikroskopisch kleinen Zellfäden besteht und Karposporophyt genannt wird. Der Karposporophyt bildet nackte, diploide, unbegeißelte Karposporen. Aus diesen keimt eine dritte, ebenfalls diploide Generation heran, die Tetrasporophyt genannt wird und wieder einen autonomen Organismus bildet. Vom Laien kann der Tetrasporophyt kaum vom Gametophyten unterschieden werden, da beide in Form und Größe fast identisch sind. Der Tetrasporophyt bildet unter Reduktionsteilung aus jeder Sporenmutterzelle vier haploide Sporen aus, die Tetrameiosporen genannt werden. Sie verdriften, und aus ihnen wächst wieder eine neue haploide Generation heran.
Einzelnachweise
- ↑ Hochspringen nach: a b
c d Lin-Yong Qiu, Alexander B. Taylor, Hilary A. McManus:
 Evolution of the life cycle in land plants.
Journal of Systematics and Evolution 50 (2012), S. 171–194.
Evolution of the life cycle in land plants.
Journal of Systematics and Evolution 50 (2012), S. 171–194.
- ↑ Joachim W. Kadereit, Christian Körner, Benedikt Kost, Uwe Sonnewald: Strasburger Lehrbuch der Pflanzenwissenschaften. Springer Spektrum, Berlin/Heidelberg 2014, S. 591.


© biancahoegel.de
Datum der letzten Änderung: Jena, den: 11.06. 2026